
This information is for educational purposes only and is neither intended to nor implied to be a substitute for professional medical advice.
Always seek the advice of your physician or other qualified health provider prior to using a dietary supplement or with questions regarding a medical condition.
Introduction
Qualia Mind was designed by taking into account key neural pathways and processes. It contains nootropic compounds, neuro-vitamins, amino acids, choline donors, and herbal adaptogens. Each ingredient has stand-alone actions; they can also have additive or synergistic actions when combined, complementing each other in support of a particular cognitive pathway or process. We invite you to read Qualia: The Complex Intelligence Behind Its Formulation and Neurohacker Dosing Principles to better understand some of the thought processes behind the formulation. In this article, we want to share how individual ingredients stack together for supporting a particular pathway (e.g., choline signaling, dopamine signaling) or process (e.g., neurogenesis, membrane health).
Neurochemistry—Choline Signaling Stack
Supporting pathways and processes that overlap with choline-related signaling is the foundation of a great nootropic stack. Without adequate levels of choline in the brain (and the ability to use it well), other parts of a stack are unlikely to work well. Key parts of our design of this stack include: (1) augment the choline pool; (2) choose choline sources and other nutrients that give full choline pathway support; (3) optimize acetylcholine turnover (i.e., recycling and breakdown); and (4) support balanced signaling and neuroprotection.
The Institute of Medicine (IOM) has classified choline as an essential nutrient. While humans can make a small amount of choline in the liver, it’s primarily provided by the diet. Choline (and its metabolites) serve three main roles in the body. One of these is neurotransmission (as acetylcholine). Another is use in cell membrane structure and signaling (as phosphatidylcholine). And the third is use in methylation pathways (as trimethylglycine).
The main fate of dietary choline is the synthesis of phosphatidylcholine. This occurs via the cytidine diphosphate-choline (CDP-choline) pathway (or Kennedy pathway), which plays a central role in choline homeostasis.1 Phosphatidylcholine accounts for around 95% of the total choline pool in most tissues. The remaining 5% includes free choline, phosphocholine, glycerophosphocholine (GPC), CDP-choline, and acetylcholine, as well as other choline-containing phospholipids.
Qualia Mind contains one form of choline—Citicholine (CDP-choline)—and another nutrient—Uridine Monophosphate—that enter the CDP-choline pathway prior to phosphatidylcholine.* In this pathway, choline is initially converted to phosphocholine by choline kinase (CK), using ATP (the energy currency of cells) as a phosphate donor. Magnesium is a cofactor for this enzyme reaction. Next comes the rate-limiting step of the pathway (i.e., the slowest step in the pathway so akin to a bottleneck). An enzyme called CTP:phosphocholine cytidylyltransferase (CCT) uses cytidine triphosphate (CTP) to convert phosphocholine into CDP-choline (also known as Citicoline). We chose to include Citicholine because it is the choline form produced after the rate-limiting step and can cross the blood-brain-barrier.*
We included Uridine Monophosphate because it plays a role in the CTP molecule used during the rate-limiting step. CTP is synthesized from uridine triphosphate, which is produced by phosphorylation of uridine monophosphate. Oral Uridine Monophosphate can support healthy brain levels of CDP-choline,2 so is an indirect (i.e., non-choline) way to augment this pathway.*
In the last step in the pathway, CDP-choline is esterified with diacylglycerol (DAG) by cholinephosphotransferase (also called choline/ethanolamine-phosphotransferase [CEPT]) to produce phosphatidylcholine. Essential fatty acids (e.g. eicosapentaenoic acid (EPA) and Docosahexaenoic Acid (DHA)) are involved in the creation of DAG.
Phosphatidylcholine is the end product of the CDP-choline pathway. Since most choline in the body and brain are in the form of phosphatidylcholine, it serves as a reservoir or pool for choline. Sunflower Lecithin is a source of phospholipids including phosphatidylcholine.
Free choline and choline metabolites can be regenerated by the controlled breakdown of phosphatidylcholine. The main routes for phosphatidylcholine breakdown are via phospholipases that hydrolyze the two acyl chains of phosphatidylcholine to yield free fatty acids and glycerylphosphorylcholine (GPC). The subsequent hydrolysis of GPC into glycerol 3-phosphate and choline is catalyzed by glycerophosphodiesterase (GDE) according to cellular needs.
Qualia Mind included another form of choline—Alpha-Glycerophosphocholine (Alpha-GPC)—as part of our goal of whole pathway support. Alpha-GPC delivers choline to the brain because it can cross the blood-brain barrier (BBB).* Choline in the Alpha-GPC form enters and augments the choline pool after phosphatidylcholine. Since enzyme reactions in the choline pathways tend to be bidirectional (i.e., they can flow in either direction), more availability of this intermediate, may allow phosphatidylcholine to be used for other purposes (rather than being broken down to augment choline pools).3,4 By supplying choline in the Alpha-GPC form, we are supplying a form of choline that can be used in the brain to form free choline, but which enters the choline pool at a different point in the pathway.*
Given the different kinetics (i.e., absorption and metabolism) and different places these ingredients interact with choline pathways, including Alpha-GPC, Citicholine and Uridine Monophosphate for choline signaling support allows the upregulation of the CDP-choline pathway and turnover of the choline/phosphatidylcholine cycle, ultimately producing a higher availability of choline for acetylcholine synthesis and other uses.* Because of their complementary actions, this combination allows us to use lower doses of each.
When we designed this stack, we also chose to include Phosphatidylserine. Phosphatidylserine is synthesized from phosphatidylcholine (and phosphatidylethanolamine, which can also originate from phosphatidylcholine).5 By including Phosphatidylserine—a phospholipid used to promote healthy cell membranes*—it may spare the use of phosphatidylcholine for its synthesis, resulting in more phosphatidylcholine availability for choline regeneration and, consequently, acetylcholine production.

Acetylcholine is a neurotransmitter (i.e., a messenger molecule released by nerve cells to signal other cells). It plays a key role in enhancing alertness (when we wake up), sustaining attention, and improving learning and memory.6 Neurons that use or are affected by acetylcholine are referred to as cholinergic. The critical acetylcholine building blocks are choline and acetyl groups.
The availabilities of both acetyl-coenzyme A (acetyl-CoA) and choline will greatly influence the brain’s ability to produce acetylcholine. Vitamin B5 (pantothenic acid) is used as a cofactor in CoA synthesis.*7 CoA is then acetylated to form acetyl-CoA, predominantly via the mitochondrial pyruvate dehydrogenase complex, and then translocated to the cytoplasm.
The pyruvate dehydrogenase complex consists of 3 enzymes that convert pyruvate (a key intermediate in several metabolic pathways) into acetyl-CoA—vitamins B1 (thiamin) and B3 (niacin) are needed for this complex. Vitamin B5 was added to this stack to support biosynthesis of CoA. Thiamine and Niacinamide were added for vitamin B1 and B3 support, respectively. In vitro evidence suggests that acetyl groups from compounds such as Acetyl-L-Carnitine or N-Acetyl-L-Tyrosine might contribute to acetylcholine synthesis.*8 Acetyl-L-Carnitine also plays a role in neuronal mitochondrial function.*
Choline is taken up by cholinergic neurons from the extracellular fluid by the high-affinity choline uptake transporter (ChT) and accumulates in the synaptic terminal (where it can be combined with acetyl molecules). The activity of ChT shows substantial plasticity and can be affected by activation of cholinergic neurons. The production of acetylcholine can be limited because of the rate of choline uptake by ChT. This step can be enhanced by Docosahexaenoic Acid (DHA), the main structural component of the neuronal cell membrane, which improves the capacity of ChT to transport choline into neurons, thereby increasing its availability for acetylcholine synthesis.9 This might be part of the reason that dietary Docosahexaenoic Acid (DHA) supports healthy acetylcholine levels.*10
Acetylcholine is produced in nerve terminals of cholinergic neurons. This synthesis process uses the enzyme choline acetyltransferase (ChAT) to catalyze the transfer of the acetyl group from acetyl-CoA to choline.3 ChAT is the rate-limiting enzyme step in acetylcholine synthesis. Increasing the availability of neuronal choline and acetyl groups is crucial to support acetylcholine synthesis. It’s also important that ChAT activity be able to keep pace with demands. Bacopa monnieri was included in this stack because it might support the activity of ChAT, and consequently, the output of acetylcholine.11
Acetylcholine exerts its effects by activating two main classes of receptors. The first type are nicotinic acetylcholine receptors (nAChR). These are ligand-gated ion channels (i.e., ionotropic; opening to let charged minerals called ions pass through cell membranes) permeable to sodium (Na+), potassium (K+), and calcium (Ca2+) ions, whose activation causes a fast depolarization and excitation. Magnesium, in its ion form Mg2+, is involved in balancing ion flows into cell membranes through the nAChR ionotropic receptors. Phosphatidylserine supports acetylcholine release and influences ionotropic acetylcholine receptor function.*12,13 The second type are muscarinic acetylcholine receptors (mAChR). These are G protein-coupled receptors (i.e., metabotropic; acting through intracellular signaling molecules called second messengers) that activate signaling pathways inside cells. Both classes of acetylcholine receptors participate in the mechanisms of memory.
In contrast to most other neurotransmitters, the synaptic action of acetylcholine is not terminated by reuptake. Instead, cleanup is done exclusively through enzymatic hydrolysis (i.e., inactivation) by acetylcholinesterase (AChE) to yield choline and acetate. The free choline can then be taken up again by the synaptic terminal and recycled to acetylcholine (i.e., starting the synthesis process again). Bacopa monnieri might help enhance the effects of acetylcholine by slowing the activity of AChE, leaving acetylcholine available to produce greater receptor stimulation.*14 Celastrus paniculatus and Coleus forskohlii might also influence the activity of AChE.*,15 Huperzine A (a nootropic compound) has improved acetylcholine levels in animal and human studies and influences cholinesterase enzyme activity.*16-19 Ginkgo biloba (a neuroadaptogen) appears to have protective and regulatory effects on aspects of acetylcholine signaling;*20,21
Cholinergic signaling might also be enhanced by other mechanisms. For example, adenosine (a sleep inducing factor) is a neuromodulator that reduces cholinergic activity. Blocking adenosine receptors with antagonist molecules can counter adenosine’s effects, supporting healthy activity of acetylcholine.22 Caffeine is a non-selective and high-affinity adenosine receptor antagonist (wakefulness promoting activity of coffee is believed to be related to this effect) and might indirectly enhance cholinergic activity.*22,23 Theobromine (a compound found in chocolate) is related to and synergistic with caffeine, and it is also an adenosine receptor antagonist (albeit with lower affinity than caffeine). It can also support healthy activity of acetylcholine, and it has a slower onset and longer duration of effects than caffeine.*23,24

Neurochemistry—Glutamate Signaling Stack
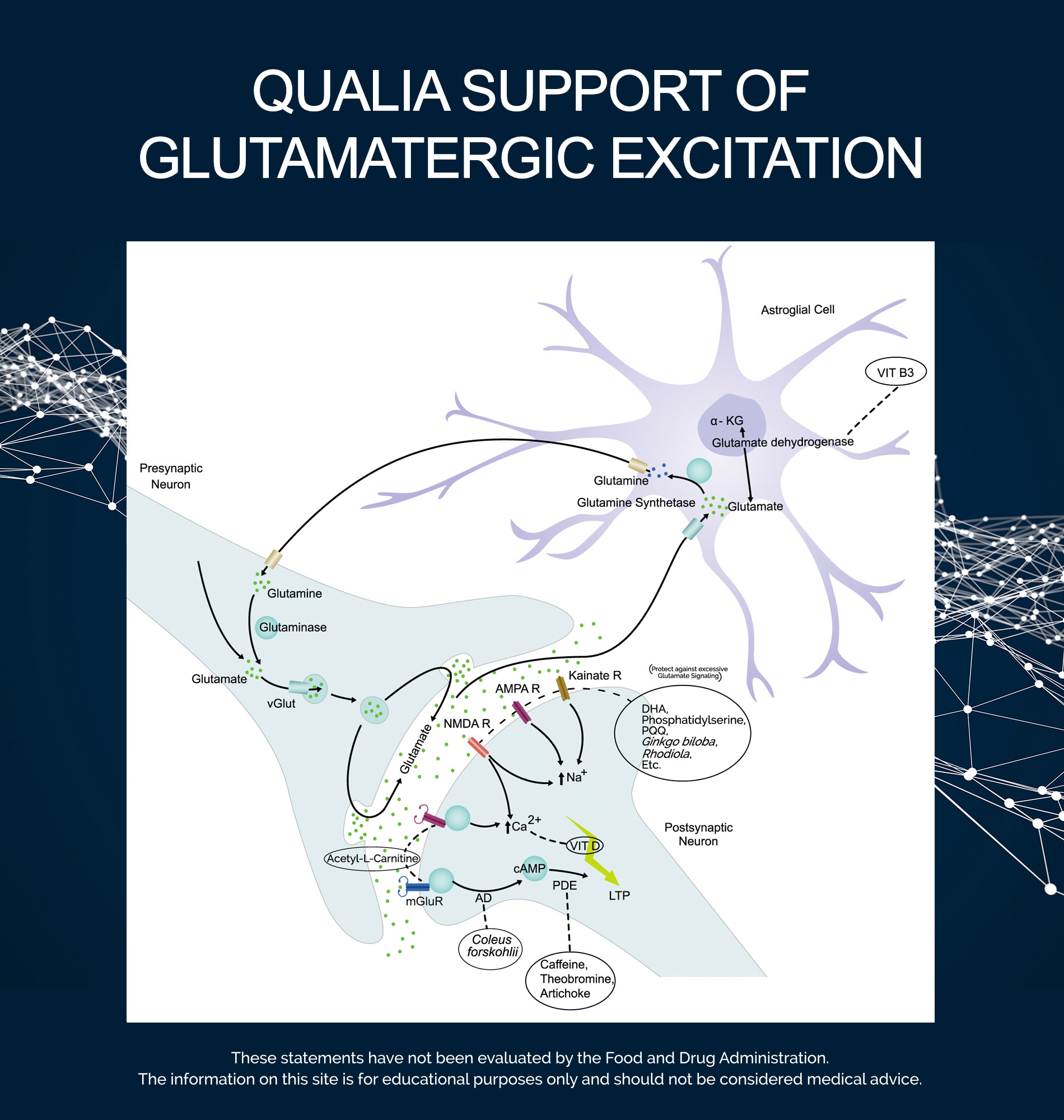
The most important design factor for a glutamate signaling stack is to promote receptor sensitivity to glutamate, while protecting against excessive glutamate signaling. Other considerations are support of (1) enzyme function involved in glutamate synthesis, signaling, and cleanup; and (2) endogenous neuroprotective systems.*
Glutamate is the most abundant neurotransmitter in our brain and central nervous system (CNS). It is involved in virtually every major excitatory brain function. While excitatory has a very specific meaning in neuroscience, in general terms, an excitatory neurotransmitter increases the likelihood that the neuron it acts upon will have an action potential (also called a nerve impulse).
Because it is the main molecule promoting neuronal excitation, glutamate is the principal mediator of cognition, emotions, sensory information, and motor coordination, and is linked to the activity of most other neurotransmitter systems. But glutamate is not a “more is better” molecule. Glutamatergic communication requires the right concentrations of glutamate be released in the right places for only small amounts of time. Less than this results in poor communication. More than this can be neurotoxic and can damage neurons and neural networks.
Glutamate, and its receptors, are central elements in memory formation and retrieval because of their role in the key cellular mechanism of memory and learning called long-term potentiation (LTP). LTP is a form of synaptic plasticity, a term that refers to the biochemical processes through which synapses respond to patterns of activity, either by strengthening in response to increased activity, or by weakening in response to decreased activity. LTP is the persistent strengthening component of plasticity. It is one of the major cellular mechanisms that underlies how the brain encodes memories.
Glutamate does not cross the blood-brain barrier and must be synthesized in neurons from precursor molecules that can get into the brain. In the brain, glutamine is the fundamental building block for glutamate. The most prevalent biosynthetic pathway synthesizes glutamate from glutamine using an enzyme called glutaminase. Glutamate can also be produced from glucose through a metabolic pathway that begins with the conversion of glucose to pyruvate (a process called glycolysis). Pyruvate then ethers the tricarboxylic acid (TCA) cycle (also called the Krebs cycle or citric acid cycle). The TCA cycle forms multiple important intermediates. One of these intermediates is α-ketoglutarate (α-KG). α-KG can be used to produce glutamate. An enzyme called glutamate dehydrogenase, which uses vitamin B3 (NAD+) as a coenzyme, is responsible for this reaction. Niacinamide has been included to support this coenzyme function. This same enzyme can reconvert glutamate back into α-KG. Because of this enzyme, glutamate and α-KG can be continuously converted into each other. This dynamic equilibrium is a key intersection between anabolic and catabolic pathways and allows the body to shift resources in whichever direction is required.
There are two general types of glutamate receptors. One type are called ionotropic receptors: Glutamate binding to these receptors allows the entry of ions (i.e., electrically charged minerals such as sodium or calcium) into the cell. There are three classes of ionotropic glutamate receptors: (1) N-methyl-D-aspartate (NMDA), (2) α-amino-3-hydroxy-5-methyl-4-isoxazole propionic acid (AMPA), and (3) kainate receptors. The second type of receptors are linked to molecules that will activate intracellular signaling pathways subsequent to glutamate binding. These are called G protein-coupled or metabotropic receptors. Metabotropic glutamate receptors (mGluR) modulate synaptic transmission (i.e. neuronal communication) by regulating the activity of a wide variety of ion channels, including ionotropic glutamate receptors, as well as receptors for other neurotransmitters. Carnitine (from Acetyl-L-Carnitine) might support and protect metabotropic glutamate receptors.*25
Glutamate plays a prominent role in neural circuits involved with synaptic plasticity and LTP via NMDA and AMPA receptor activity, and with calcium influx into postsynaptic neurons.26 Acetylcholine’s effect on memory is also associated with LTP. Cholinergic projections to the hippocampus can modulate glutamatergic signaling and LTP.27
Magnesium plays a key role in the regulation of NMDA receptor excitability. At a resting membrane potential (i.e., pre-glutamate excitation), Mg2+ ions act to block the ion channel of NMDA receptors. Glutamate excitation relieves the Mg2+ blockade and calcium (Ca2+) influx through NMDA receptors is allowed. Low levels of Mg2+ in the CNS (i.e., a relative magnesium insufficiency) contribute to the hyperexcitability of NMDA receptors. As a consequence, fewer NMDA channels will be blocked, more NMDA channels will be opened at lower membrane potentials, and an excessive calcium influx can occur at a lower threshold of glutamate signaling, leading to excitotoxicity. Neuronal Mg2+ concentrations are therefore of major neurophysiological importance. However, magnesium insufficiency is common.28 Given the important role of calcium in the mechanisms of LTP, we added Vitamin D3 to facilitate calcium absorption.29
Excessive glutamatergic activity can be toxic, an effect known as glutamate excitotoxicity. This effect is due to an excessive entry of calcium ions into the cell, which can trigger cell-damaging pathways. Therefore, it is important that the enhancement of glutamate signaling is supported in a way that avoids such effects while at the same time allowing LTP to occur. This approach to neuroprotection is achieved by blocking excessive activation of NMDA receptors, while leaving normal function relatively intact, thereby may help normalize calcium influx.30
We included several ingredients that help regulate NMDA receptor activity,* with a goal of protecting neurons from glutamate-induced toxicity by promoting balanced glutamate signaling and/or supporting receptors. Celastrus paniculatus might be neuroprotective against excitotoxicity, possibly through modulation of NMDA receptor activity.*31,32 Huperzine A appears to support balanced NDMA receptor binding and may reduce glutamate excitotoxicity by reducing glutamate-induced calcium mobilization.*33,34 Vitamin C and Pyrroloquinoline Quinone (PQQ) might support NMDA glutamate receptors, which could provide protection against excitotoxicity.*35,36 Taurine may reduce the affinity of NMDA glutamate receptors to glycine, which is needed for their activation,37 while still inducing LTP.*38
In later phases of LTP, the reinforcement of synaptic connections requires intracellular gene transcription and protein synthesis.. Calcium entry into the postsynaptic neuron activates adenylate cyclase (AD) and leads to the production of cyclic AMP (cAMP). cAMP is a second messenger. It acts as an important signaling molecule inside cells. cAMP activates signaling pathways that lead to an upregulation of mRNA translation that sustains late LTP.39 LTP is also regulated by the activity of phosphodiesterase-4 (PDE4), the enzyme that hydrolyzes cAMP.40 Both adenylate cyclase activation and phosphodiesterase inhibition have been shown to promote long-lasting LTP.41,42
cAMP is an example of a seesaw effect in a pathway. Adenylate cyclase is on one end, pushing cAMP production up, but PDE4 is on the other end, pushing it back down. Coleus forskohlii and Artichoke extract were included in the stack to synergistically target synaptic plasticity by impacting the push-pull of the cAMP pathway. Forskolin (found in Coleus forskohlii) may induce late LTP by supporting adenylate cyclase activity, leading to healthy intracellular levels of cAMP,43 whereas Artichoke extract might influence cAMP levels indirectly via effects of PDE4, further augmenting intracellular levels of cAMP.44,45 Artichoke’s effects might be further supported by Caffeine and Theobromine, which also exert effects on PDE4.46
Other ingredients in our stack that support aspects of glutamate signaling include: (1) Neuroadaptogens Ginkgo biloba and Rhodiola rosea, which may protect neurons from excessive glutamate excitation;*47-49 and (2) Neurolipid compounds Phosphatidylserine and Docosahexaenoic Acid (DHA), which are used in cell membranes and appear to support balanced glutamate receptor signaling and function.*50,51
Neurochemistry—Dopamine Signaling Stack
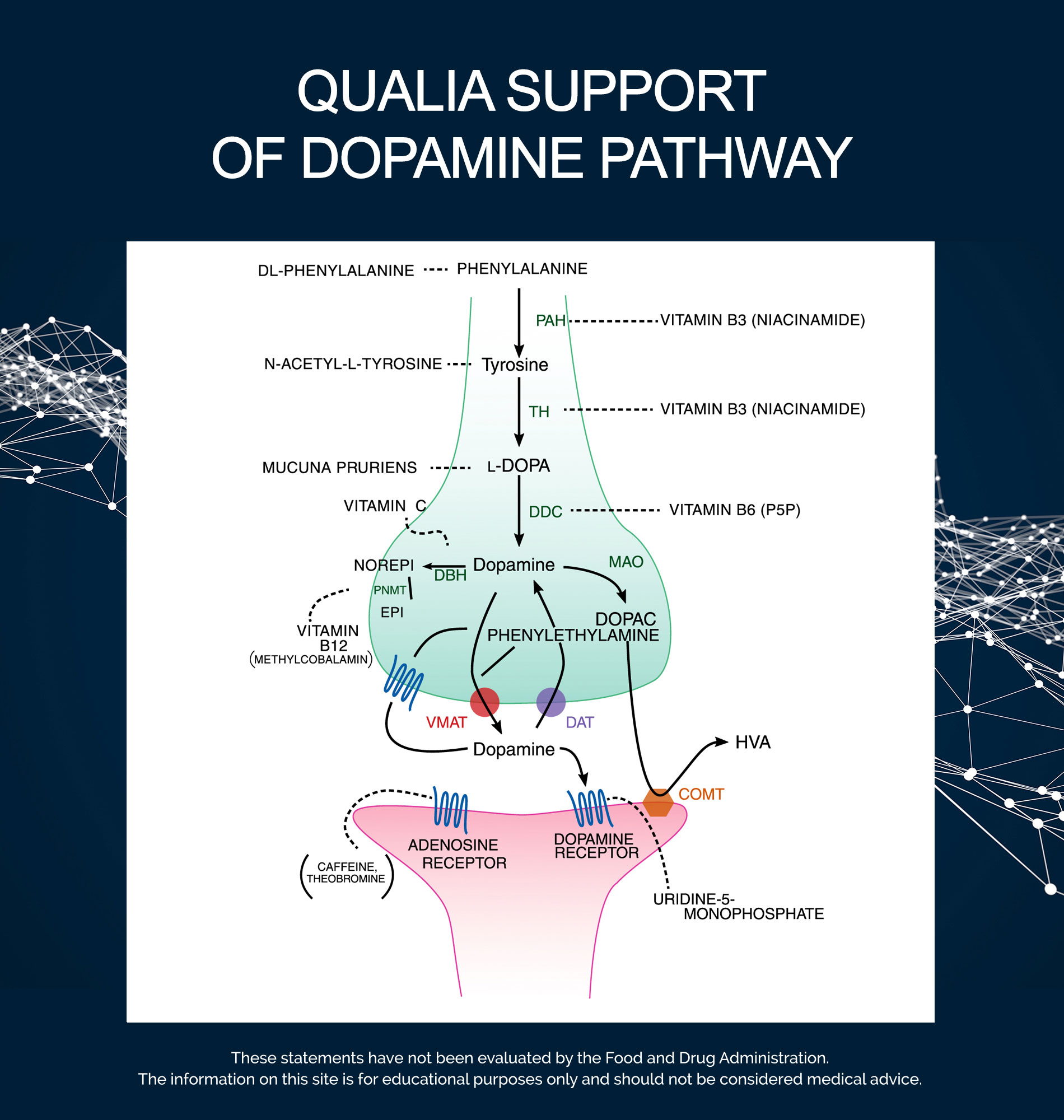
Key parts of designing a dopamine stack are: (1) augment the precursor pool of compounds used to make it; (2) give full pathway support; (3) support enzyme function involved in dopamine synthesis, signaling, and cleanup; and (4) promote balanced signaling and neuroprotection. Let’s put these pieces together now.
Dopamine is one of the main neurotransmitters in the brain. It is most commonly recognized for its role in reward, motivation, and pleasure, but also plays a crucial part in modulating focus, motivation, cognitive flexibility, and emotional resilience. In addition to these creative-productive capacities and states, dopamine is one of the main regulators of motor control and coordination of body movements.
Dopamine is synthesized both at the nerve terminals, and in the cell bodies of dopaminergic neurons. But it does not cross the blood-brain barrier and must be synthesized in the brain from precursors molecules. There are 3 key precursor molecules in the dopamine synthesis pathway that can get into the brain—L-phenylalanine, L-tyrosine, and L-DOPA. Dopamine can be made starting from any of these molecules, with the molecules entering the pathway at different points.
The most fundamental building block is the essential amino acid L-phenylalanine (supplied as DL-Phenylalanine). L-tyrosine (supplied as N-Acetyl-L-Tyrosine) is the next step in the dopamine pathway. Since it can be synthesized from L-phenylalanine by phenylalanine hydroxylase (PAH), L-tyrosine is considered conditionally essential. Under certain circumstances (e.g., illness, high stress, cognitive demands) where the body might not be able to make enough to meet demands, it becomes important to get more L-tyrosine from the diet.
The rate-limiting step in this L-DOPA pathway is the enzyme tyrosine hydroxylase (TH). This is the enzymatic step that turns L-tyrosine into L-DOPA, as the pathway progresses from L-Phenylalanine → L-Tyrosine → L-DOPA → Dopamine. Mucuna pruriens—a member of the legume (i.e., bean) family—is included in this stack because it is a natural source of L-DOPA.*52 We use this herbal ingredient to supply an amount of L-DOPA that a person would consume if they ate about 3-6 ounces of fava beans (fava beans are considered one of the richest food sources of L-DOPA). This ingredient thereby supports the production of dopamine by entering the pathway after the rate-limiting step.*
By including Mucuna pruriens, N-Acetyl-L-Tyrosine, and DL-Phenylalanine, the stack supports three different steps, with different kinetics, of the dopamine synthesis pathway, allowing for a prolonged and sustained availability of precursor resources to be recruited for its synthesis.* Importantly, by taking advantage of additive effects, we can support the dopaminergic pathway using lower doses of each ingredient.
Deficiency in any precursor amino acid or in any cofactor in the catecholaminergic anabolic pathways can impair the synthesis of all three catecholamine neurotransmitters (e.g., dopamine, epinephrine, norepinephrine). L-DOPA is converted to dopamine by aromatic-L-amino-acid decarboxylase (AAAD, also known as DOPA decarboxylase [DDC]) with Pyridoxal-5’-Phosphate (the active form of Vitamin B6) as the coenzyme.53 Both tyrosine hydroxylase (TH) and phenylalanine hydroxylase (PAH) require tetrahydrobiopterin as a coenzyme. Tetrahydrobiopterin is synthesized from guanosine triphosphate (GTP) through a NADPH-dependent pathway.54 Vitamin B3, as Niacinamide, is a precursor to NADPH,55 and can therefore indirectly support the activity of both enzymes.
Dopamine is used as precursor in the synthesis of the monoamine neurotransmitters norepinephrine and epinephrine. Dopamine is converted into norepinephrine by the enzyme dopamine β-hydroxylase, with L-ascorbic acid (Vitamin C) as cofactor; norepinephrine is then converted into epinephrine. Vitamin C is included to support the conversion of dopamine to norepinephrine by dopamine β-hydroxylase.
After synthesis, dopamine is transported into synaptic vesicles by a vesicular monoamine transporter. Once released, dopamine binds to and activates either postsynaptic receptors or presynaptic autoreceptors. When an action potential is elicited in the postsynaptic neuron, dopamine molecules are released from their receptors and retaken up into the presynaptic cell or into surrounding glial cells by the dopamine transporter (DAT) or by the plasma membrane monoamine transporter (VMAT).
Within neurons, dopamine is degraded into inactive metabolites by monoamine oxidase (MAO), which catalyzes the oxidative deamination of dopamine into DOPAL. It is then converted into DOPAC by aldehyde dehydrogenase (ALDH) and into homovanillic acid (HVA), the primary metabolite of dopamine, by catechol-O-methyltransferase (COMT). Dopamine can also be metabolized (i.e., inactivated) by COMT into 3-methoxytyramine, using SAMe as a methyl donor.
COMT is synthesized in two forms, soluble (i.e., free-floating) and membrane-bound; the latter is the primary form in the brain. COMT is dependent on magnesium. COMT-dependent extracellular dopamine inactivation is of particular importance in brain regions with low expression of the presynaptic dopamine transporter—the recycling mechanism that takes up and returns dopamine into the message sending neurons for inactivation or reuse.
Several ingredients are included because of their actions on postsynaptic receptor signaling. Uridine Monophosphate has a unique role in the dopamine stack. Although uridine may reduce the density of dopamine receptors, it seems to enhance their signal transduction and turnover rate, supporting dopamine-dependent behaviors.56 Uridine Monophosphate also enhances potassium-evoked dopamine release.57 Both Caffeine and Theobromine play a role in this stack due to their antagonism of adenosine receptors. Since adenosine receptor activation reduces dopaminergic activity, slowing activity of adenosine receptors can indirectly contribute to an enhanced dopaminergic signaling.22-24
Resilience and Adaptogenic Stack
Resilience can be thought of as a generalized capability to cope with emergent challenges or stress. This is the “adaptive” part of the science of complex adaptive systems. The body follows a type of response called hormensis when responding to certain challenges. An adaptive response implies too little or too much exposure results in suboptimal response, while a just-right range of exposure—the hormetic zone—where there's a generally favorable biological response. Exercise follows this type of response. A range of exercise amounts will produce improved fitness and resilience, but below this amount (e.g., sedentary behavior) or above (e.g., injury, overtraining) won’t. Several ingredients, including herbal adaptogens, were included in this formulation due to their resilience-enhancing effects.
Adaptogen is a term used to classify substances that support resistance to stress when dosed in the correct range. These substances typically promote homeostasis, exerting a normalizing or stabilizing effect on physiological processes.58 In a sense, adaptogens act to toughen us up, by producing a small amount of stress, which in turn allows our body to respond better to other sources of stress.*
A central element to the maintenance of homeostasis is the neuroendocrine system, a set of mechanisms by which the hypothalamus maintains homeostasis. The hypothalamic–pituitary–adrenal (HPA) axis, a complex network of interactions among the hypothalamus, the pituitary gland and the adrenal gland, is of particular interest due to its role in the regulation of stress responses, mood, and energy storage and expenditure.59
There are several important molecules whose production is induced by stress and that mediate adaptogenic effects and coordinate the adaptive stress response. These include the stress hormones cortisol, corticosterone and corticotropin-releasing factor. They also include nitric oxide, stress-activated protein kinases, and heat shock proteins (such as HSP70). These factors, and many more, are part of an innate response to stress. The result of this complex response produces improved tolerance and protection from future stress. It also promotes adaptation via feedback-mediated downregulation of the activated HPA axis.58, 60
Rhodiola rosea is one of the most potent adaptogens.* Its adaptogenic effects have been confirmed in multiple studies. It has shown stress-protective and endocrine normalizing effects. It has also been shown to improve mood, cognitive performance, and attention, and to relieve fatigue in stress-related conditions.*61 Rhodiola rosea exerts an anti-stress effect by balancing the HPA axis.*62 Rhodiola rosea has been shown to modulate HSP70 expression,63 and may reduce the levels of stress-activated protein kinases, nitric oxide, cortisol,64 corticotropin-releasing factor, and peripheral corticosterone levels.*61
Several other ingredients have adaptogenic effects. Bacopa monnieri (through the action of bacoside A) has been shown to reduce the levels of the stress marker HSP70 in the brain.*65,66 Gingko biloba may reduces stress and corticosterone levels.*67 Mucuna pruriens has been effective in reducing serum cortisol levels and stress-induced oxidative changes.*68 Phosphatidylserine might reduce the levels of adrenocorticotropin and cortisol in response to physical stress.*69
Vitamin B5 is often thought of as an “anti-stress vitamin.” It plays a role in this stack due to its importance in maintaining the integrity of the cells of the adrenal cortex.70 The adrenal glands are part of the HPA axis and are responsible for the synthesis of corticosteroids, which have important roles not only in the stress response, but also in metabolism, immune response, and behavior.
cAMP—the second messenger discussed in the choline stack—influences activity of the HPA axis. The HPA produces a variety of signaling molecules called hormones. The defining characteristic of hormones is that they are made in certain places, but are able to travel to distant parts of the body to regulate physiology and behavior. Cortisol is an example: It is made by the adrenal glands—the A from HPA—but has effects throughout the body. While hormones have distant effects, these effects occur on the outside of cells. cAMP is called a second messenger because it relays the message from these hormonal first messengers into the cell. Because of this, it is a critical player in the intracellular signal transduction mechanisms that control the secretory activity of the hypothalamus, the pituitary gland, and the adrenal glands, as well as of other endocrine organs.71 PDE4 inhibitors are known to stimulate the HPA axis.72 This is, at least in part, because PDE4 plays a pivotal role in the breakdown of cAMP. Both Coleus forskohlii (an adenylate cyclase activator43) and Artichoke extract (a PDE4 inhibitor),44,45 by increasing the levels of cAMP, can support the HPA axis.* This effect is further augmented by Caffeine and Theobromine, which also influence PDE4 activity.*46
Neurogenesis, Neuroprotection, and Neuroplasticity Stack
Neurogenesis is the process of producing new neurons (i.e., nerve cells). Neuroprotection refers to preserving the neurons we have. Neuroplasticity is the ability of the brain to change the connections between neurons (and activity levels) throughout an individual's life. The generation of new neurons, preservation of existing ones, and the creation of new connections between them is a set of interrelated processes that greatly contribute to the optimization of brain function.
Neurotrophins—nerve growth factor (NGF), brain-derived neurotrophic factor (BDNF), neurotrophin-3 (NT-3), and neurotrophin-4 (NT-4)—are a family of growth factors that regulate the development, maintenance, survival, and function of the nervous system. Neurotrophin support is an emphasis of this stack.
BDNF is of particular relevance. BDNF has been reported to regulate the proliferation of neuronal precursors and to promote the differentiation, maturation and integration of newborn neurons in the hippocampus. BDNF has also shown marked effects on dendrite and spine development and maturation, increasing dendrite branching, length, and complexity. And, BDNF is involved in synaptogenesis (i.e., formation of connections between neurons) and synaptic maturation.73–76 Several ingredients enhance the production of BDNF: Ginkgo biloba,77 Taurine,78 L-Theanine,79 and a combination of Vitamin B12 and omega-3 fatty acids (DHA and EPA).*80
NGF is also involved in the proliferation, growth, maintenance, and survival of neurons. It promotes peripheral nerve regeneration and myelin repair.81 Ingredients that promote the synthesis or the activity of NGF include Huperzine A,82 PQQ83 and Phosphatidylserine.84
Other ingredients with neuroprotective or neuroplasticity enhancing effects include: Ginkgo biloba has been shown to promote neurogenesis and neuronal survival.*77 Huperzine A has influenced proliferation of hippocampal neural stem cells.*81 Rhodiola rosea has enhanced neurogenesis and neuronal regeneration.*85,86 Uridine Monophosphate supported dendritic spines and promoted neurite outgrowth.*57,87 Bacopa monnieri enhanced neurite branching and proliferation.*11 A combination of Vitamins B1, B6 and B12 has also been shown to enhance peripheral nerve regeneration.*88
Cell Membrane Integrity Stack
The cell membrane separates the interior of all cells from the outside environment. It protects the cell from its surroundings, controls the movement of substances in and out of cells, and plays a major role in signaling (i.e., communication between what's inside and outside of cells). The cell membrane is made predominantly from specialized fats or lipids.
Our formulation includes several ingredients intended to support membrane health and function, because cell membranes, particularly neuronal membranes, are essential for cognitive performance.
When we discussed the choline signaling stack, the CDP-choline pathway was mentioned. It is one half of a bigger pathway known as the Kennedy pathway. The other half of the Kennedy pathway is the CDP-ethanolamine pathway, which produces a different phospholipid end product called phosphatidylethanolamine. A big emphasis of this stack is supporting the Kennedy pathway.
Phosphatidylserine is an essential component of cell membranes. It accounts for 13–15% of the phospholipids (i.e., specialized fat molecules that make up most of the cell membrane) in the human cerebral cortex. In the plasma membrane, phosphatidylserine is localized in the cytoplasmic layer where it’s part of protein docking sites required for the activation of several important signaling pathways. In synapses, phosphatidylserine has a significant role in modulating neurotransmitter release through exocytosis (i.e., expelling these molecules out of the cell) by influencing calcium-dependent fusion of synaptic vesicles and the plasma membrane.*5 Phosphatidylserine can also be converted to other phospholipids, particularly phosphatidylethanolamine, another important structural fat used in the cell membrane, so supports the second half of the Kennedy pathway.*
Alpha-GPC, CDP-Choline and Uridine Monophosphate contribute to membrane health by supporting the first half of the Kennedy pathway (i.e., phosphatidylcholine synthesis). Besides being the major source of choline for the choline pool, phosphatidylcholine is a primary component of cell membranes. The enhanced production of phosphatidylcholine, along with phosphatidylethanolamine via phosphatidylserine, contributes to the structural integrity of cell membranes.*
Docosahexaenoic acid (DHA) is an essential fatty acid. It is the most abundant polyunsaturated fatty acid/omega-3 fatty acid in the brain and the major polyunsaturated fatty acid of neuronal cell membranes. DHA supports healthy membrane fluidity and flexibility, which affects several membrane functions, including the release and uptake of neurotransmitters. DHA also modulates membrane transport systems, membrane bound enzymes, receptors and ion channels, thereby influencing cell signaling.89 It is therefore essential for proper functional and structural support of cell membranes.*
Uridine Monophosphate and DHA support the production of synaptic proteins and phospholipids, synaptic membranes and dendritic spines.*90,91
Nerve fibers (axons) are long projections from nerve cells to remote tissues and muscles. Many axons are surrounded by a a lipid-rich substance called a myelin sheath that acts a lot like the insulation on an electrical wire. The production of this myelin sheath is called myelination. Axonal membranes need myelin to be able to transmit information efficiently. Myelination is improved by Vitamin D3.92
Cell Energy and Metabolism Support Stack
The body’s (and brain’s) "energy currency" is called adenosine triphosphate (ATP). Most ATP is produced in specialized structures within cells called mitochondria. A central set of reactions involved in ATP production are collectively known as the citric acid cycle, or Krebs cycle. Given the brain’s high metabolic rate, it is a big user of this energy currency. ATP is also used throughout the body for cell energy production and metabolism.
The overall ATP production process is called aerobic respiration. This process starts with the generation of pyruvate from the simple sugar glucose—a process called glycolysis—in the liquid found inside cells (i.e., cytosol). The pyruvate is then transported into mitochondria, where it is converted into acetyl-CoA. Acetyl-CoA is a pivotal molecule in cellular metabolism because it donates the main substrate of the citric acid cycle, the acetyl group. In the citric acid cycle (also known as the tricarboxylic or Krebs cycle), a series of chemical reactions generate NADH and FADH2, which in turn transfer the energy generated by the citric acid cycle to the electron transport chain, where ATP is produced.93
B vitamins play a key role in many steps during the ATP production process.94 Because B vitamins are involved in the different interrelated pathways of energy generation, deficiency or inadequacy in any of the B vitamins can have a negative impact on the whole process (i.e., cell energy production suffers). We included several vitamins because of the major roles they play. Vitamin B1 (used in the mitochondrial pyruvate dehydrogenase complex as an example), Vitamin B3 (needed to produce NAD+ and NADH) and Vitamin B5 (required for acetyl-CoA formation) are essential in mitochondrial aerobic respiration due to their roles as coenzymes in the citric acid cycle and the electron transport chain, and, consequently, in the production of ATP.*93 [Note: Coenzymes are parts of certain enzymes. Many of the coenzymes are derived from vitamins.]
Besides being a central molecule in metabolism and energy production, Acetyl-CoA is also involved in many other enzymatic processes. Acetyl-CoA is estimated to be a cofactor for about 4% of all known enzymes.70 The addition of Vitamin B5 to this stack is therefore meant to support cellular energy production by providing the precursor to Acetyl-CoA, and to augment performance of enzymes at a more system-wide level.* The inclusion of Acetyl-L-Carnitine and N-Acetyl-Tyrosine provides acetyl groups to augment the acetyl pool for the synthesis of acetyl-CoA.*
Vitamin B6 (in its coenzyme or active form of Pyridoxal-5′-Phosphate [P5P]) is used in a wide variety of enzymatic reactions. P5P is a coenzyme in many metabolic pathways important for cellular energy generation, including glycogen (a complex sugar) breakdown and amino acid metabolism.93
Other ingredients in this stack also support cellular metabolic pathways in several ways. Vitamin B12, as Methylcobalamin, is a biologically active form of cobalamin (i.e., a coenzyme form) needed for activity of the methionine synthase enzyme. Methionine synthase plays an important role in cellular metabolic processes associated with the transfer of methyl groups (one-carbon metabolism). In the methionine synthase reaction, Methylcobalamin accepts and donates a methyl group. This allows for the conversion of methyltetrahydrofolate (an active form of folic acid) and homocysteine into tetrahydrofolate and methionine. Methionine is then further metabolized into S-adenosylmethionine (SAMe), a universal methyl donor. Because of its role in the regeneration of SAMe, which is used extensively in biology as a source of active methyl groups, Vitamin B12 sits at the crossroads of a pathway that impacts synthesis of many metabolically important compounds, including purines, pyrimidines, and amino acids.93
After donating its acetyl group, N-Acetyl-Tyrosine yields the amino acid L-tyrosine, a building block for proteins. Likewise, DL-Phenylalanine includes the L-phenylalanine form used for protein assembly.
Uridine Monophosphate is primarily recognized as one of the building blocks of RNA, but it is also involved in other important biochemical processes. It can be converted into uridine diphosphate glucose (UDP-glucose), which promotes brain glycogen synthesis. It is also used in the CDP-choline pathway as a precursor of the CTP molecule needed for the synthesis of membrane phospholipids.*95
Acetyl-L-Carnitine is converted into carnitine, which plays a key role in the transport of long-chain fatty acids into mitochondria for ATP generation.* Vitamin C contributes to healthy carnitine levels, because it is a cofactor in two enzymes that are necessary for its synthesis.96
Pyrroloquinoline quinone (PQQ) has an important role in supporting cellular energy generation, primarily via its action on mitochondria. PQQ has been linked to the activation and expression of molecules with important roles in cellular energy metabolism and mitochondrial biogenesis (process cells use to produce more mitochondria to support greater ATP production).97,98 Prolonged PQQ intake has been shown to support mitochondrial biogenesis, and improve mitochondria efficiency, by acting as a growth factor.*98,99
Calcium is important in the regulation of cellular energy production. It’s involved in regulation of the citric acid cycle, due to the sensitivity of some of this cycle’s enzymes to the mitochondrial levels of calcium.100 The included Vitamin B5 in its calcium salt form (Calcium Pantothenate) acts as a minor calcium donor to augment calcium pools. We further supported calcium availability by adding Vitamin D3, which facilitates calcium absorption from the diet.29
cAMP has been mentioned previously. It plays important roles in many areas of metabolism as an intracellular second messenger. It is produced by the adenylate cyclase enzyme, and broken down by phosphodiesterase enzymes including PDE4. PDE4 inhibitors indirectly stimulate cellular metabolism by increasing cAMP levels, which stimulates the breakdown of fats (a process called lipolysis) and activates protein kinase A, which in turn phosphorylates (i.e., modulates) several enzymes involved in glucose and lipid metabolism.101 Caffeine, Theobromine and Artichoke extract interact with cellular metabolism through their PDE4-inhibitory activity.*44–46 By activating adenylate cyclase, leading to healthy intracellular levels of cAMP, Coleus forskohlii also contributes to this effect.*43
Cerebral Antioxidant Defense Stack
Reactive Oxygen Species (ROS) are a normal byproduct of healthy metabolism—they are created by the oxidation and reduction (redox) reactions used by mitochondria when ATP is produced. ROS were once thought of as unwanted byproducts of mitochondria energy generation (i.e., the free-radical theory of aging). Newer understandings have led to a more nuanced mitohormesis understanding of ROS. This is the mitochondrial equivalent of the hormesis principle, previously mentioned with the Resilience and Adaptogenic Stack.
Mitohormesis implies that a zone of ROS causes mitochondria to toughen up, build adaptive capacities, and perform better. It also implies that excessive ROS can damage cellular membranes and impair cell energy processes (including ATP generation). The body has evolved a complex system of antioxidant defenses to allow it to adapt to and protect itself from ROS. A molecule called glutathione (and its related enzymes) plays a central role in antioxidant defense. Other antioxidant enzymes, such as catalase and superoxide dismutase, are also critical parts of the overall antioxidant defenses.
The brain, due to its high metabolic rate, consumes 20% of the total body oxygen, making it a big producer of ROS. Excessive ROS production, and the oxidative damage this can cause, is thought to be one of the main contributing agents to premature brain aging and cognitive decline. Therefore, it’s important that the brain’s antioxidant defense system be supported in ways that allow it to deal efficiently with ROS and maintain redox balance. Several ingredients in this formulation act directly in this area, either as antioxidants, by promoting antioxidant defenses, or both.*
Vitamin C is a potent antioxidant. As an electron donor, it can reduce, and thereby neutralize ROS.102 Vitamin C is essential for the maintenance of the redox balance of the brain.*103
Pyrroloquinoline quinone (PQQ) is a redox cofactor that reduces oxidants and is continuously recycled into its active form by glutathione.*104 PQQ is 100–1000 times more efficient in redox cycling than other quinone biofactors and is capable of continuously performing repeated oxidation and reduction reactions.*105
Vitamin D3 has antioxidant properties.* It inhibits lipid peroxidation,106 suggesting that it protects cellular membranes (which have a high lipid content) from oxidation.* Vitamin D is also associated with major plasma redox systems.107 L-Theanine has antioxidant effects and protects neurons from damage and hypoxia (circumstances characterized by insufficient oxygen availability).*108
Several of the included herbal extracts have antioxidant properties due to their phytochemical components, such as flavonoid compounds, which are among the most effective exogenous antioxidant molecules. Celastrus paniculatus may reduce lipid peroxidation and free radical generation. It has shown free-radical-scavenging activity for the superoxide anion and hydroxyl radical, and it reduces H2O2-induced cytotoxicity and DNA damage. It also supports the antioxidant molecule glutathione, and antioxidant enzymes catalase and superoxide dismutase.*109–111 Bacopa monnieri modulates the levels of superoxide dismutase, and protects against oxidative damage from metals in the brain.*66 Rhodiola rosea is neuroprotective against toxins and oxidative stress. It also supports the production of antioxidant enzymes.*112 Ginkgo biloba has antioxidant effects and preserves mitochondrial function, which is important for maintaining the redox balance of the cell.*113 Mucuna pruriens reduces the levels of free radicals, ROS, and lipid peroxidation, while supporting the activity of superoxide dismutase and catalase antioxidant enzymes, and Vitamin C levels in stressed individuals.*68,114 Coleus forskohlii exhibits significant antioxidant and ROS scavenging activities.*115 Artichoke extract has helped support healthier response to oxidative stress.*116
Cerebral Blood Flow Support and Blood-Brain Barrier Penetration Stack
Optimal cerebral blood flow is of obvious importance, not only because it is fundamental for the proper delivery of oxygen and metabolic ingredients, but also for the removal of waste products of cellular metabolism from the brain. Cerebral blood flow is also fundamental for the delivery of the ingredients included in this formulation. And, it’s important to choose compounds and forms of nutrients that can pass into the brain (i.e., gain access through the blood-brain barrier). Some of our ingredients were chosen based on these considerations.
Nitric oxide (NO) is a biological messenger and signaling molecule. It plays an important role in circulation. NO is used by endothelial cells—cells that line the interior surface of blood and lymph vessel—as a messenger that instructs the smooth muscle surrounding blood vessels to relax, causing the widening of blood vessels (vasodilation) and increasing blood flow. NO is produced by enzymes called nitric oxide synthases (endothelial NOS, eNOS, in this case) in a reaction that requires NADPH (vitamin B3-dependent). This stack includes two ingredients that can indirectly support NO’s blood flow improving effects. Vitamin C stimulates endothelial NO synthesis.*117 Vitamin B3 (Niacinamide) is a precursor to NADPH.*55
Vitamin C plays an important role in the synthesis of collagen and elastin,*118 important structural components of the blood vessels. Elastin confers elasticity and collagen confers strength, both essential for blood flow and general vascular health. Niacinamide provides blood flow benefits by increasing microvascular density and microcirculation.*119,120
Gingko biloba promotes cerebral blood flow. It has several active compounds that improve circulation, reduce clot formation, strengthen the walls of capillaries, and protect nerve cells from low oxygen conditions.*121
One of the criteria for our formulation design was to choose ingredients known to reach and have effects in the brain (i.e., be capable of penetrating the blood-brain barrier).* In general, fat-soluble and smaller molecules are more able to penetrate. Bigger compounds often need help (i.e., specific transporter proteins). Phosphatidylserine, a lipid molecule with evidence showing support of human cognitive function, appears to follow the fat-soluble rule of thumb—it crosses the BBB.*122
While the above is a general rule of thumb, there can be exceptions. Docosahexaenoic acid (DHA) is a fat-soluble essential fatty acid. Its synthesis is extremely limited within the brain, but it is important for brain structure and function.* DHA appears to require the help of specific transporter proteins for access to the brain via the BBB.123 Feeding DHA significantly unregulated these transport proteins in an animal study, and supported healthy brain DHA levels.*124
Some of our ingredients are included in forms that are known to enhance their bioavailability and/or their ability to cross the BBB. For example, Acetyl-L-Carnitine, the acetylated version of L-carnitine, can cross the BBB and provide better cognitive benefits.*125 N-Acetyl-L-Tyrosine is a more bioavailable version of the dopamine precursor L-tyrosine.*126 Oral Thiamine can support free thiamine levels in the brain.*127 Citicoline and Alpha-GPC are forms of choline that can cross the blood-brain barrier.*
Additional Supportive Mechanisms
Coffee, and the caffeine it contains, are alertness promoting. They can have a variety of benefits for alertness-related performance. L-Theanine was included to synergize with and reduce caffeine-induced jitteriness and vasoconstriction.*128 Caffeine and L-Theanine are commonly stacked because of their well-reported synergistic effect in enhancing cognition, mood, vigilance and attention.129,130 Bacopa Monnieri has anxiolytic effects without sedation. Therefore, one of the purposes of its inclusion was also to reduce jitteriness in order to balance our stack.*131
Other added benefits from the ingredients in this formulation include:
Healthy Inflammation Response from Coleus forskolli,132 DHA,133 and Celastrus paniculatus.*134
Analgesic effects from Bacopa monnieri,135 Celastrus paniculatus136 and D-Phenylalanine (included in the racemic mixture DL-Phenylalanine).*137,138
Reduced fatigue from Coleus forskolli,139 Rhodiola rosea140 and PQQ.*141
Support sleep quality from Taurine142 and PQQ.*141
Gastroprotective effects from Celastrus paniculatus.*143
Support aerobic capacity from Phosphatidylserine.*144
*These statements have not been evaluated by the Food and Drug Administration (FDA). This product is not intended to diagnose, treat, cure, or prevent any disease.
References
Li Z, Vance DE. J Lipid Res. 2008;49(6):1187-1194. doi:10.1194/jlr.R700019-JLR200.
Cansev M, Watkins CJ, Van Der Beek EM, Wurtman RJ. Brain Res. 2005;1058(1-2):101-108. doi:10.1016/j.brainres.2005.07.054.
Fagone P, Jackowski S. Biochim Biophys Acta - Mol Cell Biol Lipids. 2013;1831(3):523-532. doi:10.1016/j.bbalip.2012.09.009.
Gibellini F, Smith TK. UBMB Life. 2010;62(6):414-428. doi:10.1002/iub.337.
Kim H-Y, Huang BX, Spector AA. Prog Lipid Res. 2014;56:1-18. doi:10.1016/j.plipres.2014.06.002.
Hasselmo ME. Curr Opin Neurobiol. 2006;16(6):710-715. doi:10.1016/j.conb.2006.09.002.
Tahiliani AG, Beinlich CJ. Vitam Horm. 1991;46:165-228. http://www.ncbi.nlm.nih.gov/pubmed/1746161.
Doležal V, Tuček S. J Neurochem. 1981; 36(4):1323-1330. doi:10.1111/j.1471-4159.1981.tb00569.x.
Hyman BT, Spector AA. J Neurochem. 1982;38(3):650-656. http://www.ncbi.nlm.nih.gov/pubmed/7057184.
Minami M, et al. Pharmacol Biochem Behav. 1997; 58(4):1123-1129.
Aguiar S, Borowski T. Rejuvenation Res. 2013;16(4):313-326. doi:10.1089/rej.2013.1431.
Suzuki S, et al. J Nutr. 2001; 131(11):2951-2956. doi:10.1093/jn/131.11.2951.
Casamenti F, et al. Eur J Pharmacol. 1991; 194(1):11-16.
Bhanumathy M, Harish MS, Shivaprasad HN, Sushma G. Pharm Biol. 2010;48(3):324-327. doi:10.3109/13880200903127391.
Yang Q-R, Wu H-Z, Wang X-M, Zou G-A, Liu Y-W. J Asian Nat Prod Res. 2006;8(4):355-360. doi:10.1080/10286020500172236.
Wang G, et al. Nan Fang Yi Ke Da Xue Xue Bao. 2006; 26(11):1660-1662.
Liang Y, Tang X. Acta Pharmacol Sin. 2006; 27(9):1127-1136. doi:10.1111/j.1745-7254.2006.00411.x.
Liang YQ, et al. Cell Mol Neurobiol. 2008; 28(1):87-101. doi:10.1007/s10571-007-9158-9.
Zhu S-Z, et al. Neurosci Lett. 2016; 631:70-78. doi:10.1016/j.neulet.2016.07.009.
Stein C, et al. J Pharm Pharm Sci. 2015; 18(4):634-646. doi:26626253.
Kehr J, et al. Int psychogeriatrics. 2012; 24 Suppl 1:S25-34. doi:10.1017/S1041610212000567.
Fredholm BB. Pharmacol Toxicol. 1995;76(2):93-101. doi:10.1111/j.1600-0773.1995.tb00111.x.
Burnstock G. Adv Exp Med Biol. 2013;986:1-12. doi:10.1007/978-94-007-4719-7-1.
Franco R, Oñatibia-Astibia A, Martínez-Pinilla E. Nutrients. 2013;5(10):4159-4173. doi:10.3390/nu5104159.
Llansola M, et al. Metab Brain Dis. 2002;17(4):389-397.
Carlson NR, Birkett MA. Learning and memory. In: Physiology of Behavior. 12th edition. Pearson Education Limited; 2017.
Ge S, Dani JA. J Neurosci. 2005;25(26):6084-6091. doi:10.1523/JNEUROSCI.0542-05.2005.
Rosanoff A, et al. Nutr Rev. 2012; 70(3):153-164. doi:10.1111/j.1753-4887.2011.00465.x.
Christakos S, Dhawan P, Porta A, Mady LJ, Seth T. Mol Cell Endocrinol. 2011;347(1-2):25-29. doi:10.1016/j.mce.2011.05.038.
Lipton SA. Nat Rev Drug Discov. 2006;5(2):160-170. doi:10.1038/nrd1958.
Godkar PB, Gordon RK, Ravindran A, Doctor BP. J Ethnopharmacol. 2004;93(2-3):213-219. doi:10.1016/j.jep.2004.03.051.
Godkar PB, Gordon RK, Ravindran A, Doctor BP. Phytomedicine. 2006;13(1-2):29-36. doi:10.1016/j.phymed.2003.11.011.
Zhang JM, Hu GY. Neuroscience. 2001;105(3):663-669. http://www.ncbi.nlm.nih.gov/pubmed/11516831.
Ved HS, Koenig ML, Dave JR, Doctor BP. Neuroreport. 1997;8(4):963-968. http://www.ncbi.nlm.nih.gov/pubmed/9141073.
Majewska MD, Bell JA. Neuroreport. 1(3-4):194-196. http://www.ncbi.nlm.nih.gov/pubmed/1983355.
Aizenman E, Hartnett KA, Zhong C, Gallop PM, Rosenberg PA. J Neurosci. 1992;12(6):2362-2369. http://www.ncbi.nlm.nih.gov/pubmed/1318959.
Chan CY, Sun HS, Shah SM, et al. Adv Exp Med Biol. 2013;775:45-52. doi:10.1007/978-1-4614-6130-2_4.
del Olmo N, Suarez LM, Orensanz LM, et al. Eur J Neurosci. 2004;19(7):1875-1886. doi:10.1111/j.1460-9568.2004.03309.x.
Wong ST, Athos J, Figueroa XA, et al. Neuron. 1999;23(4):787-798. http://www.ncbi.nlm.nih.gov/pubmed/10482244.
Ahmed T, Frey JU. Neuroscience. 2003;117(3):627-638. http://www.ncbi.nlm.nih.gov/pubmed/12617967.
Navakkode S, Sajikumar S, Frey JU. J Neurosci. 2004;24(35):7740-7744. doi:10.1523/JNEUROSCI.1796-04.2004.
Barad M, Bourtchouladze R, Winder DG, Golan H, Kandel E. Proc Natl Acad Sci USA. 1998;95(25):15020-15025. http://www.ncbi.nlm.nih.gov/pubmed/9844008.
Gobert D, Topolnik L, Azzi M, et al. J Neurochem. 2008;106(3):1160-1174. doi:10.1111/j.1471-4159.2008.05470.x.
Xu B, Li X-X, He G-R, et al. Eur J Pharmacol. 2010;627(1-3):99-105. doi:10.1016/j.ejphar.2009.10.038.
Röhrig T, Pacjuk O, Hernández-Huguet S, Körner J, Scherer K, Richling E. Med (Basel, Switzerland). 2017;4(4). doi:10.3390/medicines4040080.
Francis SH, Sekhar KR, Ke H, Corbin JD. Handb Exp Pharmacol. 2011;(200):93-133. doi: 10.1007/978-3-642-13443-2_4.
Zhu L, et al. Zhongguo Yao Li Xue Bao. 1997; 18(4):344-347.
Cho KS, et al. Phytother Res. 2016; 30(1):58-65. doi:10.1002/ptr.5500.
Palumbo DR, et al. Phytother Res. 2012; 26(6):878-883. doi:10.1002/ptr.3662.
Gagné J, et al. Brain Res. 1996; 740(1-2):337-345.
Wang X, et al. Neuroreport. 2003; 14(18):2457-2461. doi:10.1097/01.wnr.0000093483.65585.86.
Pulikkalpura H, Kurup R, Mathew PJ, Baby S. Sci Rep. 2015;5(1):11078. doi:10.1038/srep11078.
Montioli R, Cellini B, Dindo M, Oppici E, Voltattorni CB. Biomed Res Int. 2013;2013:1-10. doi:10.1155/2013/161456.
Thöny B, Auerbach G, Blau N. Biochem J. 2000;347 Pt 1:1-16. http://www.ncbi.nlm.nih.gov/pubmed/10727395.
Belenky P, Bogan KL, Brenner C. Trends Biochem Sci. 2007;32(1):12-19. doi:10.1016/j.tibs.2006.11.006.
Farabegoli C, Pich EM, Cimino M, Agnati LF, Fuxe K. Acta Physiol Scand. 1988;132(2):209-216. doi:10.1111/j.1748-1716.1988.tb08319.x.
Wang L, Pooler AM, Albrecht MA, Wurtman RJ. J Mol Neurosci. 2005;27(1):137-145. doi:10.1385/JMN:27:1:137.
Panossian A. Ann N Y Acad Sci. 2017;1401(1):49-64. doi:10.1111/nyas.13399.
Smith SM, Vale WW. Dialogues Clin Neurosci. 2006;8(4):383-395. http://www.ncbi.nlm.nih.gov/pubmed/17290797.
Munck A, Guyre PM, Holbrook NJ. Endocr Rev. 1984;5(1):25-44. doi:10.1210/edrv-5-1-25.
Panossian A, Wikman G, Sarris J. Phytomedicine. 2010;17(7):481-493. doi:10.1016/j.phymed.2010.02.002.
Xia N, Li J, Wang H, Wang J, Wang Y. Exp Ther Med. 2016;11(1):353-359. doi:10.3892/etm.2015.2882.
Hernández-Santana A, Pérez-López V, Zubeldia JM, Jiménez-del-Rio M. Phytother Res. 2014;28(4):623-628. doi:10.1002/ptr.5046.
Panossian A, Hambardzumyan M, Hovhanissyan A, Wikman G. Drug Target Insights. 2007;2:39-54. http://www.ncbi.nlm.nih.gov/pubmed/21901061.
Anbarasi K, Kathirvel G, Vani G, Jayaraman G, Shyamala Devi CS. Neuroscience. 2006;138(4):1127-1135. doi:10.1016/j.neuroscience.2005.11.029.
Kar Chowdhuri D, Parmar D, Kakkar P, Shukla R, Seth PK, Srimal RC. Phyther Res. 2002;16(7):639-645. doi:10.1002/ptr.1023.
Rapin JR, Lamproglou I, Drieu K, Defeudis F V. Gen Pharmacol. 1994;25(5):1009-1016. doi:10.1016/0306-3623(94)90111-2.
Shukla KK, Mahdi AA, Ahmad MK, Jaiswar SP, Shankwar SN, Tiwari SC. Evidence-based Complement Altern Med. 2010;7(1):137-144. doi:10.1093/ecam/nem171.
Monteleone P, Beinat L, Tanzillo C, Maj M, Kemali D. Neuroendocrinology. 1990;52(3):243-248. doi:10.1159/000125593.
Sweetman L. Pantothenic acid. In: Coates PM, Betz JM, Blackman MR, et al., eds. Encyclopedia of Dietary Supplements. 2nd edition. Informa Healthcare; 2010.
Sette C, Vicini E, Conti M. Mol Cell Endocrinol. 1994;100(1-2):75-79. doi:10.1016/0303-7207(94)90282-8.
Kumari M, Cover PO, Poyser RH, Buckingham JC. Br J Pharmacol. 1997;121(3):459-468. doi:10.1038/sj.bjp.0701158.
Waterhouse EG, An JJ, Orefice LL, et al. J Neurosci. 2012;32(41):14318-14330. doi:10.1523/JNEUROSCI.0709-12.2012.
Chan JP, Cordeira J, Calderon GA, Iyer LK, Rios M. Mol Cell Neurosci. 2008;39(3):372-383. doi:10.1016/j.mcn.2008.07.017.
Gao X, Smith GM, Chen J. Exp Neurol. 2009;215(1):178-190. doi:10.1016/j.expneurol.2008.10.009.
Wang L, Chang X, She L, Xu D, Huang W, Poo M. J Neurosci. 2015;35(22):8384-8393. doi:10.1523/JNEUROSCI.4682-14.2015.
Tchantchou F, Lacor PN, Cao Z, et al. J Alzheimers Dis. 2009;18(4):787-798. doi:10.3233/JAD-2009-1189.
Caletti G, Almeida FB, Agnes G, Nin MS, Barros HMT, Gomez R. Behav Brain Res. 2015;283:11-15. doi:10.1016/j.bbr.2015.01.018.
Wakabayashi C, Numakawa T, Ninomiya M, Chiba S, Kunugi H. Psychopharmacology (Berl). 2012;219(4):1099-1109. doi:10.1007/s00213-011-2440-z.
Rathod RS, Khaire AA, Kale AA, Joshi SR. Biochimie. 2016;128-129:201-208. doi:10.1016/j.biochi.2016.08.009.
Aloe L, Rocco ML, Balzamino BO, Micera A. Curr Neuropharmacol. 2015;13(3):294-303. http://www.ncbi.nlm.nih.gov/pubmed/26411962.
Ma T, Gong K, Yan Y, et al. Brain Res. 2013;1506:35-43. doi:10.1016/j.brainres.2013.02.026.
Yamaguchi K, Sasano A, Urakami T, Tsuji T, Kondo K. Biosci Biotechnol Biochem. 1993;57(7):1231-1233. doi:10.1271/bbb.57.1231.
De Simone R, Ajmone-Cat MA, Tirassa P, Minghetti L. J Neuropathol Exp Neurol. 2003;62(2):208-216. doi:10.1093/jnen/62.2.208.
Qu QZ, Zhou Y, Zeng YS, et al. PLoS One. 2012;7(1):e29641. doi:10.1371/journal.pone.0029641.
Sheng Q-S, Wang Z-J, Zhang J, Zhang Y-G. Neuroreport. 2013;24(5):217-223. doi:10.1097/WNR.0b013e32835eb867.
Sakamoto T, Cansev M, Wurtman RJ. Brain Res. 2007;1182:50-59. doi:10.1016/j.brainres.2007.08.089.
Becker KW, Kienecker EW, Dick P. Neurochirurgia (Stuttg). 1990;33(4):113-121. doi:10.1055/s-2008-1053569.
Hashimoto M, Hossain S, Al Mamun A, Matsuzaki K, Arai H. Crit Rev Biotechnol. 2017;37(5):579-597. doi:10.1080/07388551.2016.1207153.
Cansev M, Ulus IH, Wang L, Maher TJ, Wurtman RJ. Neurosci Res. 2008;62(3):206-209. doi:10.1016/j.neures.2008.07.005.
Wurtman RJ, Ulus IH, Cansev M, Watkins CJ, Wang L, Marzloff G. Brain Res. 2006;1088(1):83-92. doi:10.1016/j.brainres.2006.03.019.
Chabas J-F, Stephan D, Marqueste T, et al. PLoS One. 2013;8(5):e65034. doi:10.1371/journal.pone.0065034.
Berg J, Tymoczko J, Stryer L, eds. Biochemistry. 7th edition. W.H. Freeman and Company; 2011.
Kennedy D. B. Nutrients. 2016;8(2):68. doi:10.3390/nu8020068.
Cansev M. Brain Res Rev. 2006;52(2):389-397. doi:10.1016/j.brainresrev.2006.05.001.
Rebouche CJ. Am J Clin Nutr. 1991;54(6 Suppl):1147S-1152S. http://www.ncbi.nlm.nih.gov/pubmed/1962562.
Bauerly K, Harris C, Chowanadisai W, et al. PLoS One. 2011;6(7):e21779. doi:10.1371/journal.pone.0021779.
Chowanadisai W, Bauerly KA, Tchaparian E, Wong A, Cortopassi GA, Rucker RB. J Biol Chem. 2010;285(1):142-152. doi:10.1074/jbc.M109.030130.
Harris CB, Chowanadisai W, Mishchuk DO, Satre MA, Slupsky CM, Rucker RB. J Nutr Biochem. 2013;24(12):2076-2084. doi:10.1016/j.jnutbio.2013.07.008.
Wan B, LaNoue KF, Cheung JY, Scaduto RC. J Biol Chem. 1989;264(23):13430-13439. http://www.ncbi.nlm.nih.gov/pubmed/2503501.
Cappelletti S, Daria P, Sani G, Aromatario M. Curr Neuropharmacol. 2015;13(1):71-88. doi:10.2174/1570159X13666141210215655.
Padayatty SJ, Katz A, Wang Y, et al. J Am Coll Nutr. 2003;22(1):18-35. doi:10.1080/07315724.2003.10719272.
Figueroa-Méndez R, Rivas-Arancibia S. Front Physiol. 2015;6. doi:10.3389/fphys.2015.00397.
Mukai K, Ouchi A, Nakano M. J Agric Food Chem. 2011;59(5):1705-1712. doi:10.1021/jf104420y.
Stites TE, Mitchell AE, Rucker RB. J Nutr. 2000;130(4):719-727. doi:10.1093/jn/130.4.719.
Wiseman H. FEBS Lett. 1993;326(1-3):285-288. http://www.ncbi.nlm.nih.gov/pubmed/8325381.
Alvarez JA, Chowdhury R, Jones DP, et al. Clin Endocrinol (Oxf). 2014;81(3):458-466. doi:10.1111/cen.12449.
Sumathi T, Asha D, Nagarajan G, Sreenivas A, Nivedha R. Environ Toxicol Pharmacol. 2016;42:99-117. doi:10.1016/j.etap.2016.01.008.
Godkar P, Gordon RK, Ravindran A, Doctor BP. Fitoterapia. 2003;74(7-8):658-669. doi:10.1016/S0367-326X(03)00190-4.
Russo A, Izzo AA, Cardile V, Borrelli F, Vanella A. Phytomedicine. 2001;8(2):125-132. doi:10.1078/0944-7113-00021.
Kumar KH, Venuprasad MP, Jayashree G V, et al. Cytotechnology. 2015;67(6):955-967. doi:10.1007/s10616-014-9733-0.
Qu ZQ, Zhou Y, Zeng YS, Li Y, Chung P. Biomed Environ Sci. 2009;22(4):318-326. doi:10.1016/S0895-3988(09)60062-3.
Eckert A, Keil U, Kressmann S, et al. Pharmacopsychiatry. 2003;36 Suppl 1(S 1):15-23. doi:10.1055/s-2003-40449.
Lampariello LR, Cortelazzo A, Guerranti R, Sticozzi C, Valacchi G. J Tradit Complement Med. 2012;2(4):331-339. doi:10.1016/S2225-4110(16)30119-5.
Bajpai VK, Park Y-H, Agrawal P. Front Life Sci. 2015;8(2):139-147. doi:10.1080/21553769.2014.998777.
Magielse J, Verlaet A, Breynaert A, et al. Mol Nutr Food Res. 2014;58(1):211-215. doi:10.1002/mnfr.201300282.
Heller R, Unbehaun A, Schellenberg B, Mayer B, Werner-Felmayer G, Werner ER. J Biol Chem. 2001;276(1):40-47. doi:10.1074/jbc.M004392200.
Harrison FE, May JM. Free Radic Biol Med. 2009;46(6):719-730. doi:10.1016/j.freeradbiomed.2008.12.018.
Smith YR, Klitzman B, Ellis MN, Kull FC. J Surg Res. 1989;47(5):465-469. http://www.ncbi.nlm.nih.gov/pubmed/2530402.
Kelleher DK, Vaupel PW. Int J Radiat Oncol Biol Phys. 1993;26(1):95-102. http://www.ncbi.nlm.nih.gov/pubmed/8482636.
Singh B, Kaur P, Gopichand, Singh RD, Ahuja PS. Fitoterapia. 2008;79(6):401-418. doi:10.1016/j.fitote.2008.05.007.
Glade MJ, Smith K. Nutrition. 2015 Jun;31(6):781-6. doi: 10.1016/j.nut.2014.10.014.
Chouinard-Watkins R, Lacombe RJS, Bazinet RP. Curr Opin Clin Nutr Metab Care. 2018 Mar;21(2):71-77. doi: 10.1097/MCO.0000000000000440.
Pan Y, Morris ER, Scanlon MJ, et al. J Neurochem. 2018 Mar 27. doi: 10.1111/jnc.14342.
Nałecz K. Mol Aspects Med. 2004;25(5-6):551-567. doi:10.1016/j.mam.2004.06.001.
Topall G, Laborit H. J Pharm Pharmacol. 1989;41(11):789-791. doi:10.1111/j.2042-7158.1989.tb06368.x.
Vignisse J, Sambon M, Gorlova A, et al. Mol Cell Neurosci. 2017;82:126-136. doi: 10.1016/j.mcn.2017.05.005.
Dodd FL, Kennedy DO, Riby LM, Haskell-Ramsay CF. Psychopharmacology (Berl). 2015;232(14):2563-2576. doi:10.1007/s00213-015-3895-0.
Owen GN, Parnell H, De Bruin EA, Rycroft JA. Nutr Neurosci. 2008;11(4):193-198. doi:10.1179/147683008X301513.
Kahathuduwa CN, Dassanayake TL, Amarakoon AMT, Weerasinghe VS. Nutr Neurosci. 2017;20(6):369-377. doi:10.1080/1028415X.2016.1144845.
Bhattacharya SK, Ghosal S. Phytomedicine. 1998;5(2):77-82. doi:10.1016/S0944-7113(98)80001-9.
Menon D, Latha K. Pharmacogn J. 2011;3(23):75-79. doi:10.5530/pj.2011.23.11.
Bazan NG, Molina MF, Gordon WC. Annu Rev Nutr. 2011;31(1):321-351. doi:10.1146/annurev.nutr.012809.104635.
Ahmad F, Khan RA, Rasheed S. J Ethnopharmacol. 1994;42(3):193-198. doi:10.1016/0378-8741(94)90085-X.
Bhaskar M, Jagtap AG. Int J Ayurveda Res. 2011;2(1):2-7. doi:10.4103/0974-7788.83173.
Kulkarni YA, Agarwal S, Garud MS.J Ayurveda Integr Med. 2015;6(2):82-88. doi:10.4103/0975-9476.146540.
Russell AL, McCarty MF. Med Hypotheses. 2000;55(4):283-288. doi:10.1054/mehy.1999.1031.
Ehrenpreis S. Acupunct Electrother Res. 1982;7(2-3):157-172. http://www.ncbi.nlm.nih.gov/pubmed/6128872.
Henderson S, Magu B, Rasmussen C, et al. J Int Soc Sports Nutr. 2005;2(2):54. doi:10.1186/1550-2783-2-2-54.
Kelly GS. Altern Med Rev. 2001;6(3):293-302. http://www.ncbi.nlm.nih.gov/pubmed/11410073.
Nakano M, Yamamoto T, Okamura H, Tsuda A, Kowatari Y. Funct Foods Heal Dis. 2012;2(8):307-324. http://functionalfoodscenter.net/files/56592277.pdf.
Kuriyama K, Hashimoto T. Adv Exp Med Biol. 1998;442:329-37. doi:10.1007/978-1-4899-0117-0_41.
Palle S, Kanakalatha A, Kavitha CN. J Diet Suppl. August 2017:1-13. doi:10.1080/19390211.2017.1349231.
Kingsley MI, Miller M, Kilduff LP, McEneny J, Benton D. Med Sci Sports Exerc. 2006;38(1):64-71. doi:10.1249/01.mss.0000183195.10867.d0.






No Comments Yet
Sign in or Register to Comment