
Viruses are everywhere cellular life is present, often in unfathomable numbers. They mutate very often, frequently by recombining with other viruses. This means that new viruses are constantly being generated.
Viruses are specific for certain types of hosts, meaning that there are bacterial viruses, plant viruses, animal viruses, etc. Therefore, fortunately, not all viruses are able to infect human cells. Still, our body is constantly being exposed to and having to deal with new viruses. Most often, they are quickly eliminated by our immune system; they may even go by unnoticed. But as the COVID-19 pandemic has left abundantly clear, every now and then, a new virus can wreak havoc on our lives.
As we’ll learn in this article, viruses are very simple, but despite their simplicity, they are very effective and impressive little creatures. We’ll also learn how our immune system rises to the challenge.
What is a Virus?
Viruses are a unique group of infectious agents. Viruses aren’t microbes because they aren’t cells. And because they aren’t cells, they aren’t typical life forms or “organisms.” [Historically, definitions of life asserted that a living organism must be composed of one or more cells, but some modern criteria allow for non-cellular life forms, of which viruses are the prime example.]
A virus is an acellular structure with a very simple structural organization: a genetic element surrounded by protein. Viruses lack metabolism and all the machinery that actual cells have. Therefore, viruses are obligate intracellular parasites that require a living host cell to replicate; outside cells, viruses are inactive. The viral genome encodes the functions needed for the virus to multiply, using the cell’s machinery to do so. [1,2]
Viruses excel at reproduction, but individual viruses don't have the machinery to build new viruses themselves. They invade cells and hijack cellular genetic tools to reproduce.
The viral genome, in other words, the DNA or RNA (nucleic acids), is the actual virus. The complete viral particle, which is the extracellular form that allows the virus to travel from one host cell to another, is called a virion. In other words, the virion is a virus (a nucleic acid) enclosed within a protein shell, called the capsid. [For the sake of simplicity, we’ll sacrifice accuracy and use the term virus even when we refer to the complete particle.]
Most bacterial viruses are naked, with no further layers, but many animal viruses have an outer lipoprotein membrane surrounding the nucleocapsid (i.e., the capsid with the nucleic acid) called the envelope. Some envelope proteins may project from the envelope surface as spikes. The capsid and the envelope protect the viral genome when the virus is outside the host cell. They are also important for the attachment of a virus to the cell it is invading, the host cell.
Viral genomes are small but diverse; they can consist of either DNA or RNA (or both DNA and RNA, but at different stages of their “life” cycle), be either single-stranded or double-stranded, be either linear or circular, for example. The largest viral genomes are DNA genomes with only a few hundred genes; the smallest viral genomes are those of some small RNA viruses that infect animals that have only two (!) genes (for scale, the smallest bacterial genome has around 100 genes).
In some viruses, the viral particle carries one or more virus-specific enzymes that are required for infection and replication, as for example, enzymes that can make RNA from an RNA template, which are needed for RNA viruses but are not available in host cells. But viruses only carry this extra baggage when they really need it.
Viruses vary widely in shapes and sizes, but they are much smaller than bacteria (although megaviruses comparable in size to some bacteria have been found). Interestingly, they are always highly symmetric structures—it’s a matter of assembly efficiency, as we’ll see. Even in viruses with complex structures, their several parts still display their own symmetrical shape. [1,2]
While each type of virus is unique, they are organized into larger families based on the shared characteristics they have with other types of viruses (i.e., enveloped vs. non-enveloped, RNA vs. DNA, single-stranded vs. double-stranded, etc.).
Viruses are designed for efficiency: a nucleic acid surrounded by a protective protein layer organized in an easily assemblable symmetrical structure.
Viral Infection and Replication Cycle
A virus cannot replicate on its own because it lacks the cellular apparatus that would allow it to do so. Therefore, a virus must invade a living cell in order to use its replication machinery. Although viral replication has some specificities depending on their host, their structure, and the type of nucleic acid they carry, all viruses go through generally similar processes in the course of their replication cycle. [1,2]
The viral replication cycle can be divided into five steps: 1) attachment to the host cell; 2) entry into the host cell; 3) synthesis of viral nucleic acid and proteins; 4) particle assembly and packaging of viral genomes; 5) release of new viruses from the cell. The period between the attachment of the virus to the host cell and the release of new virions is called the latent period of infection. At the end of this period, new viruses are released, ready to infect other cells and restart this cycle in each newly infected cell.
1) Attachment to the host cell
To gain access to an animal cell, the virus particle first needs to attach to its surface. Attachment to the host cell is achieved through the interaction between molecules on the surface of the viral particle and specific molecules on the surface of the host cell. The host molecules to which the viral particle attaches are called receptors and they are just normal cell surface molecules, with normal cellular functions, that the virus recognizes; these may be proteins, carbohydrates, glycoproteins, lipids, lipoproteins, or other cell surface structures. The virus can only attach to a cell and infect it if its specific receptor is expressed by that cell (in an unaltered form; if the receptor is altered by mutation, for example, the host may become resistant to infection by that virus). Viruses usually go for host receptors that are important for normal host cell function and that are therefore always present on the surface of the host cell.
Therefore, the type of cells that a given virus can infect is determined by the presence of a suitable receptor that the virus can recognize and to which it can attach. Therefore, attachment is a major factor in host specificity of a virus. In animals, cells in different tissues or organs often express different proteins on their cell surfaces. Consequently, viruses that infect animals often infect only certain tissues.
2) Entry into the host cell
In animal cells, the entire viral particle invades the host cell (in bacterial viruses only the viral genome and viral enzymes enter the host cell). Attachment of a virus causes changes to the host cell surface that allow the entry of the viral nucleic acid into the cell. Entry of an animal virus into a host cell generally occurs by one of three modes: fusion of the viral envelope with the host cell’s plasma membrane, entry by endocytosis (i.e., through the formation of a vesicle), or release of viral nucleic acid into the cytoplasm of the host cell. Upon entry into the cell, some viral particles lose their envelope and capsid and release their genetic material into the cytosol in a process called uncoating. There are also viruses that remain encapsidated.
3) Synthesis of viral nucleic acid and proteins
Once inside, the virus takes control of cellular functions and redirects the host cell’s metabolism to support its replication and the assembly of new viruses. Because of the genetic diversity of viruses, this stage of the viral replication cycle differs significantly among viruses.
In animals, if the viral genome is double-stranded DNA (the same as animal genomes), the host cell will have everything the virus needs to replicate; the viral genome will pass through the nuclear membrane to the nucleus and replicate using the host machinery.
RNA viruses, on the other hand, have specific needs that host cells can’t serve. Animal host cells do not have the enzymes that can make RNA from an RNA template. Therefore, RNA viruses carry in their nucleocapsid the enzymes needed to synthesize RNA, produce viral-specific messenger RNA (mRNA), and replicate the viral RNA genome.
Retroviruses, which are unusual RNA animal viruses that replicate via DNA intermediates, also carry the enzyme that can make DNA from an RNA template (called reverse transcriptase) because that’s another process host cells cannot carry out.
4) Assembly of the viral particle and packaging of viral genomes
Once the viral genome has been copied and coating proteins have been produced by the host’s machinery, the viral nucleic acid copies are packaged within their capsid and envelope (when present).
Some viruses may require assistance from host cell proteins for proper folding and assembly of their proteins and capsids, particularly those whose particles have multiple components that require multiple assembly lines functioning independently and converging in later steps to complete the final structure. But for many (if not most) viruses, the information required for the assembly of viral proteins into capsids is embedded within the amino acid sequence of the viral proteins themselves. In other words, the structural properties of viral proteins make virion assembly a spontaneous process; this is called self-assembly and it’s an extraordinary economy of resources. This is why viral particles are symmetric.
Some viruses carry out the synthesis and assembly steps within the host’s cytoplasm. To protect these processes from host defenses, some viruses reorganize membranes of cellular organelles and form membrane structures that enclose and protect their replication and assembly called viral replication complexes. Other viruses carry out synthesis and assembly in defined and concentrated areas within the cytoplasm called viroplasms. Viral replication complexes and viroplasms are also referred to as virus factories.
Enveloping typically occurs when the particle exits the animal cell, through membrane budding, during which the virus picks up part of the cell’s cytoplasmic membrane to use it as part of its envelope.
5) Release of new viruses from the cell
Eventually, the host cell ruptures and new viruses are released; the host cell dies and each new virus is free to repeat the process in a new host cell. The rupture (lysis) of the host cell is the most common outcome of a viral infection; this is called a virulent (or lytic) infection. Viruses can also be released by budding, as is common with enveloped viruses. When viral particles are released by budding, the host cell may survive and continue releasing viruses for some time.
But there are also other possible outcomes in an animal virus infection. In a latent infection, the viral DNA is integrated into the host’s genome and no new viruses are produced; host cells remain unharmed until an event triggers the virulent pathway. There are also persistent infections in which the production and release of new viruses may take place very slowly and continuously, without rupture or death of the host cells. Another possibility observed with certain animal viruses is the conversion by the virus of a normal cell into a tumor cell, a process called cellular transformation. [1,2]
Viral replication creates and releases new viral particles ready to infect other cells, start a new replication cycle, and propagate the infection.
Antiviral Immune Responses
The immune system is the collection of cells and molecules that mediate our immunity. It protects us by monitoring our body and responding to the threats it encounters, be it infectious microbes and viruses or cancer cells, for example. [3–5]
Antiviral immune defense mechanisms act to block the replication of viruses and to kill virus-infected cells, thus eliminating the sites of virus accumulation. Both innate and adaptive immune mechanisms contribute to the elimination of viral infections.
Check out our overviews of the innate immune system and the adaptive immune system to learn more about them.
The Innate Immune System Is the First Responder
Innate immune responses are the first line of defense against viruses. Innate immunity is maintained by physical and chemical barriers, soluble effector molecules in the blood and extracellular fluids, immune cells, and chemical messengers in the blood and tissues. Many viral infections can be eliminated through the action of these innate immune defenses. [3–5]
One of the main mechanisms of innate immunity against viruses involves signaling by a type of cytokines—signaling molecules with important roles in inflammation and in immune cell communication and activity—called type I interferons (IFNs). Type I IFNs include IFN-α and IFN-β, the most important in viral defense. Infection by many viruses is associated with the production of these signaling molecules whose main activity is the activation of antiviral responses.
When infected cells and cells of the immune system detect the presence of viral molecules through specific cell receptors, a cell signaling pathway is activated that leads to the production and secretion of type I IFNs. In an attempt to stop the spread of the infection, type I IFNs secreted by infected cells act on neighboring uninfected cells to inhibit viral replication. They do so by signaling through their receptors to activate the transcription of genes that increase cells’ resistance to viral infection. Type I IFNs induce the expression of enzymes that block viral replication and place uninfected cells in an antiviral state that blocks the progression of the viral infection.
Another important innate immune response to viral infections is the activity of natural killer (NK) cells. NK cells are activated by activating ligands on the surface of infected cells and by type I IFNs (and other molecules) secreted by infected cells and by other cells of the immune system. Dendritic cells, for example, produce large amounts of type I IFNs and other signaling molecules that activate NK cells when they detect the presence of viruses. The binding of type I IFNs to NK cells increases their lytic and cytotoxic activity, i.e., their ability to destroy cells. In addition to activating them, these signals make NK cells more effective in killing virally infected cells.
Natural killer (NK) cells are cells of the innate immune system whose main function is to kill infected cells. NK cells do so by releasing cytotoxic protein granules adjacently to the infected cells. These granules contain pore-forming proteins that create a route for granule enzymes to enter into infected cells. These enzymes (called granzymes) will degrade proteins within the cells and initiate signaling events that will culminate in the death of the infected cells. Killing of virus-infected cells by NK cells is an important early response to viral infection, acting before adaptive immune responses have had the time to develop.
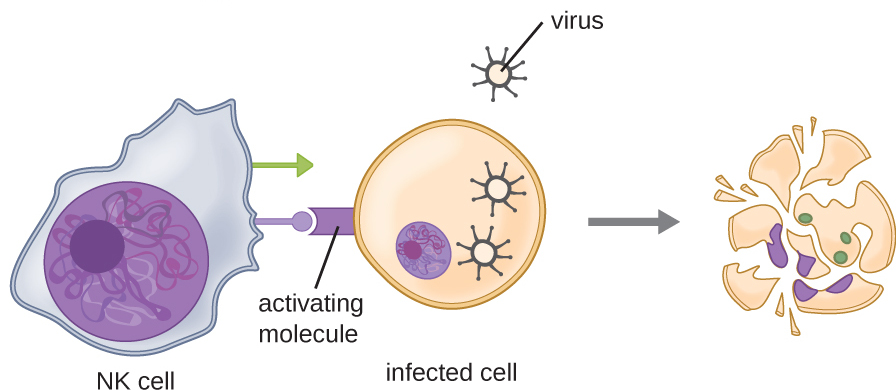
Figure 3: Destruction of an infected cell by NK cells.
Source: OpenStax, Microbiology; 17.3 Cellular Defenses. License CC BY 4.0.
The innate immune system responds to viruses by blocking viral replication and killing infected cells. Dendritic cells and NK cells are two of the immune system cell types involved in this early response against viruses.
Adaptive Immunity Is the Second Wave of Protection
When innate immune responses are insufficient to eliminate viral infections, the adaptive immune system steps in to help. Adaptive immunity is mediated by cells called lymphocytes. There are two major populations of lymphocytes called B cells and T cells involved in different types of adaptive immunity. B cells participate in humoral immunity mediated by antibodies. T cells participate in cell-mediated immunity, the branch of adaptive immunity that destroys intracellular pathogens such as viruses. [3–5]
T cells recognize antigens (the name given to a molecule that induces an immune response) that are presented at the surface of infected cells. Viruses will preferentially activate a specific type of T cells called CD8+ cytotoxic T lymphocytes (CTLs). Another type of T cells called CD4+ helper T cells respond preferentially to extracellular pathogens, but they also contribute to antiviral defense. A type of CD4+ helper T cells called Th1 cells produce a number of cytokines that contribute to antiviral defenses by stimulating the activation of CTL precursors, CD8+ CTLs, and NK cells.
The actions of CTLs are directed at killing virus-infected cells and releasing molecules that amplify the immune response. Similarly to NK cells (which are the cytotoxic cells of the innate immune system), CD8+ CTLs carry cytotoxic granules that they release in the vicinity of infected cells. Their granules also contain pore-forming proteins and protein-degrading enzymes that trigger the death of the infected cells.
Type I IFNs are also important in these adaptive immune responses. Type I IFNs increase the presentation of viral antigens on the cell surface, thereby increasing the probability that virally infected cells will be recognized and killed by CD8+ CTLs. Type I IFNs also stimulate adaptive immune responses by increasing the cytotoxicity of CD8+ CTLs.
The adaptive immune system responds to viral infections primarily through the cytotoxic T cell specialized in killing infected cells.
How We Become Immune to a Virus
The adaptive immune system has the capacity to “memorize” the infections it has come across. This capacity is possible because of the characteristics of lymphocyte responses. Lymphocytes recognize antigens through receptors present at their cell surface. Lymphocytes express millions of different types of antigen receptors. Each type is able to detect antigens with a very high degree of specificity. When a given virus infects our cells, the lymphocytes that are activated are lymphocytes that carry a receptor that is specific for that virus. These lymphocytes then proliferate to generate many copies of those specific lymphocytes. [3–5]
Some of the lymphocytes that are activated during an infection differentiate into a type of cells called memory cells. Memory B and T cells persist and circulate in the human body in a functionally quiescent or slowly cycling state for months or years after the infection is eliminated. Because they carry that antigen receptor which recognizes that virus, they retain their specificity; they “remember” the virus that activated them.
If they encounter that same antigen again, they mount a faster, greater, and more effective response. Memory B cells will generate a quick and robust antibody-mediated immune response, and memory T cells will devise quick cytotoxic responses. Because of this immune system memory, most likely, we won’t even know we were exposed to an infection again. We will have become immune to that specific threat.
We become immune to pathogens because the immune system creates a memory of an infection that allows it to respond more effectively to subsequent invasions by that specific pathogen.
Immune system memory is also the principle that underlies vaccines. [1,2] A vaccine is a biological substance capable of eliciting a mild immune response that creates immunity in our body and that protects it from future infections with the same agent. A vaccine is usually made with a weakened or inactivated form of the pathogen, or with one of its surface proteins, for example. The immune system will recognize its antigens as a threat and destroy the virus (the same is valid to other types of pathogens). As part of the immune response, memory cells will be created. If they ever come across that same antigen in the future, memory cells will mount a quick and effective response to neutralize the virus. We will be immune.
References
[1] M.T. Madigan, K.S. Bender, D.H. Buckley, W. Matthew Sattley, D.A. Stahl, Brock Biology of Microorganisms, Global Edition, Pearson Education Limited, 2019.
[2] J. Willey, L. Sherwood, C. Woolverton, Prescott’s Microbiology: Tenth Edition, McGraw-Hill Higher Education, 2017.
[3] A.K. Abbas, A.H.H. Lichtman, S. Pillai, Cellular and Molecular Immunology E-Book, Elsevier Health Sciences, 2017.
[4] K.M. Murphy, C. Weaver, Janeway’s Immunobiology: Ninth International Student Edition, W.W. Norton & Company, 2016.
[5] J. Punt, S. Stranford, P. Jones, J. Owen, Kuby Immunology, Macmillan Learning, 2018.




No Comments Yet
Sign in or Register to Comment