
Cellular senescence is a physiological process through which, in response to stress, cells stop dividing but don’t die. Some senescent cells linger indefinitely in tissues by upregulating pro-survival mechanisms called senescent cell anti-apoptotic pathways (SCAPs) and downregulating pro-apoptotic pathways [1–3]. Accumulation of lingering senescent cells occurs with aging and can impair tissue function [4]. This connection to unhealthy aging occurs in part because senescent cells produce chemical mediators, collectively known as senescence-associated secretory profile (SASP), that may disrupt tissue function and immune responses and affect health [5,6].
Maintaining balanced senescence is important because, although senescent cells can have undesirable actions, cellular senescence is also part of healthy tissue function. Some senescent cells exert beneficial effects by promoting tissue repair and regeneration and stimulating immune responses that help to maintain tissue homeostasis [1,7]. In normal conditions, these transient senescent cells are quickly eliminated by the immune system, but when their clearance fails, they can linger in tissues and gradually accumulate, becoming detrimental [1,5,6].
The accumulation of lingering senescent cells and the continued action of the hundreds of types of SASP molecules they secrete—cytokines, chemokines, matrix metalloproteinases, and other bioactive molecules—can contribute to tissue dysfunction by influencing physiological signaling pathways that disrupt immune function and promote tissue health deterioration and functional decline [1,5,6]. These tissue changes brought about by senescent cells can influence our overall health and contribute to the aging process.
Senolytics are substances that target senescent cells, normalize their overactive pro-survival mechanisms, and allow them to finally complete the process of cell death, thereby eliminating them and stopping the production of SASP mediators [4]. By doing so, senolytics support the body in protecting itself against senescent cell burden and maintaining balanced levels of cellular senescence.
Senolytics and Aging
Cellular senescence is one of the hallmarks of aging [8]. The number of senescent cells in the body increases with aging and their accumulation accelerates as we age [6,9]. The word senescence actually derives from the Latin word senex, which means senior or of old age.
The word senescence actually derives from the Latin word senex, which means senior or of old age.
Unchecked cellular senescence can be regarded as both a consequence and a cause of aging. Senescent cells build up in tissues as a consequence of several age-related changes: 1) an increase in several cellular stressors that induce senescence; 2) poorer efficiency of the immune system at finding and clearing senescent cells; and 3) a reinforcement and propagation of senescence caused by SASP factors released by lingering senescent cells [3,10].
These processes create a snowball effect of senescence build-up. SASP molecules secreted by senescent cells contribute to persistent immune signaling changes that interfere with tissue repair and regeneration and hinder healthy tissue function [6,9]. This causes progressive tissue degeneration and functional decline, age‐related dysfunction, poorer physical function, and other detrimental physiological changes that can accelerate the aging process and contribute to poorer health as we age [11–14].
Senolytics can target senescent cells and promote their elimination by normalizing their pro-survival and anti-apoptotic mechanisms and allowing them to finally complete the process of cell death, thereby also mitigating the detrimental action of SASP mediators [4]. Preclinical research has shown that the selective elimination or disruption of senescent cells with senolytics can restore healthy functions in a number of different tissues and organs, and enhance healthspan and longevity in animals [15–21].
Preclinical studies have demonstrated the ability of senolytic compounds to promote healthy metabolic function [22], mitigate age-related dysfunctions in the liver [23], support heart and kidney tissue repair after injury [18,19], counter age-related bone loss [17], and support cognitive functions in animal models of brain aging [20,21], among other beneficial actions.
Human studies with senolytics are still limited because this is a relatively new area of research. Nevertheless, the first clinical trials with senolytics had promising findings, showing the ability of senolytics to reduce markers of senescence in blood, skin, and adipose tissue in individuals with metabolic and kidney dysfunction [24] and to support physical function in a small group of individuals with lung dysfunction [25]. Many other ongoing clinical trials are assessing the potential of senolytics in the support of general health with aging.
In addition to senolytics, another approach to the management of senescent cells is SASP neutralization using senomorphics [26]. Senomorphics are compounds that neutralize or prevent the production of SASP mediators by blocking signaling pathways, disrupting the secretion of SASP molecules, or inhibiting the activity of individual SASP mediators. However, this approach can be complex because senescent cells from different tissues can produce different SASP factors and promote tissue dysfunction through different mechanisms [26]. The advantage of senolytics is that they can also abolish the production of SASP molecules, but as a consequence of targeting and eliminating the senescent cells that secrete them, which is a more permanent approach [4].
Can You Get Senolytics in Food?
Senolytics are a relatively new area of research. It was only in 2015 that the term senolytic was coined, when a study revealed two compounds that were able to eliminate senescent cells and extend healthspan in old mice [27]. As research on senolytics advances, synthetic senolytics are being developed, but importantly, many natural compounds with senolytic action are being revealed.
Many of these natural senolytics are compounds that are commonly consumed as part of the human diet. Nutritional senolytics are a growing area of research because they are a promising intervention to support healthy aging through nutrition [28].
However, although there are many different fruits, vegetables, herbs, spices, and other types of foods in the diet that contain senolytic or senomorphic compounds, no food source has been shown to contain senolytics in sufficient amounts for actual senolytic benefits, so far. In other words, in research studies, these compounds showed senolytic action (i.e., they were able to promote the death of senescent cells) only at amounts that are much higher than those available in foods.
Although there are many different fruits, vegetables, herbs, spices, and other types of foods in the diet that contain senolytic or senomorphic compounds, no food source has been shown to contain senolytics in sufficient amounts for actual senolytic benefits.
Many biological actions of natural compounds are said to be dose-dependent, meaning they only occur above specific amounts. That’s the case with the senolytic compounds identified so far: the amounts present in foods are not sufficient to achieve the dose threshold for a senolytic action. Therefore, you shouldn’t expect a senolytic benefit from those foods. There are a few examples that illustrate this well.
Quercetin was one of those first two compounds—the first natural compound—to be described as a senolytic in 2015 [27]. Quercetin is a flavonol from the flavonoid group of polyphenols (we’ll learn more about polyphenols below). Quercetin is widely distributed in fruits and vegetables. The highest concentrations of quercetin are found in onions, asparagus, capers, leafy greens such as kale and spinach, herbs such as fennel leaves, dill, and oregano, and berries such as blueberry, cherry, and cranberry, and in small quantities in many other fruits and vegetables [29,30].
A clinical trial that used a combination of senolytics that included quercetin to reduce senescence used a dose of 1000 mg of quercetin. Considering that onions, a good and common source of quercetin, provide 45 mg of quercetin per 100 g [29], you would have to eat over 2 kg of onions—close to 5 pounds—to get a serving of 1000 mg of quercetin. Alternatively, you would have to eat the outer skin and the thin hairy roots at the bottom—the parts people usually throw out—because that’s where the highest levels of quercetin in onions are found (we’ll learn why when we discuss polyphenols below) [31]. Onion skins and the tougher outer layers may have up to 48-fold more quercetin than the edible portion [32]. But you would still have to eat a lot of onion skins.
Another famous senolytic discovered not long after quercetin is fisetin. Like quercetin, fisetin is a flavonol polyphenol. Fisetin was found to be the most potent senolytic among a panel of 10 flavonoid polyphenols (that included quercetin) screened for senolytic activity in senescent mouse and human fibroblasts (i.e., the cells that produce the extracellular matrix that provides the structural frame of tissues) [33]. Fisetin is found in many dietary fruits and vegetables, including strawberries, apples, persimmon, lotus root, onions, grapes, kiwi, peach, and cucumber [34].
To get 1400 mg fisetin (the amount studied to experience senolytic benefits) from strawberries, you would have to eat 8.75 kg of strawberries—around 19 pounds!
But fisetin is also found only in very low amounts, even lower than quercetin. The richest known food source of fisetin are strawberries, but they provide only 16 mg of fisetin per 100 grams of fruit [35]. The amount of fisetin being used in ongoing clinical studies is 20 mg per kg body weight daily (orally for two consecutive days), which corresponds to a daily serving of 1400 mg for a person weighing 70 kg, i.e., around 155 pounds. To get a senolytic serving of 1400 mg fisetin from strawberries, you would have to eat 8.75 kg of strawberries—around 19 pounds!
[You can learn more about fisetin and how it compares to quercetin in our Longevity Ingredient Spotlight: Fisetin Benefits.]
The discovery of quercetin’s senolytic actions spiked interest in finding other natural compounds that could have similar properties. The search for natural senolytics revealed several other compounds such as luteolin, curcumin, piperlongumine, and oleuropein [33,36,37]. Like quercetin and fisetin, these are also present in foods in small amounts. Furthermore, they’re often found in the highest amounts in foods you use only in small quantities like herbs and spices.
Luteolin is a plant flavone, another type of flavonoid polyphenol. Luteolin is found in many fruits, vegetables, and herbs, mostly in the leaves and flowering parts. Luteolin is found in high amounts in Chinese celery (leaf celery), celery seeds, Mexican oregano, fenugreek seed, radicchio, and chamomile flowers (used to make chamomile tea); it is also found in peppers, parsley, thyme, sage, and black olives, for example [30,38].
Curcumin is a polyphenol found in turmeric that is responsible for many of its health benefits [39]. Piperlongumine is an amide-alkaloid found in several types of peppers, most abundantly in long pepper (Piper longum), from which it got its name [40]. Oleuropein belongs to a specific group of polyphenols called secoiridoids. It is present in olives, but it is found in larger amounts in olive leaves [41].
In addition to nutritional senolytics, research on natural compounds that may support the management of senescent cells has also revealed a number of nutritional senomorphics, including resveratrol, kaempferol, apigenin, and epigallocatechin gallate (EGCG).
Resveratrol is a stilbene polyphenol famously found in grape skins and red wine, but also present in peanuts, cocoa, and berries such as blueberries, bilberries, and cranberries [42–44]. Kaempferol is a flavonol found in vegetables, spices, and herbs such as kale, arugula, watercress, capers, saffron, dill, chives, and tarragon [29,30].
Apigenin is one of the most common flavones in foods. It is found in many vegetables and fruits, including parsley, Chinese celery, celery seeds, kumquat, artichoke, rutabaga, sorghum, and flax. It is also found in very high amounts in the flowers used to make chamomile tea [30,45,46]. EGCG belongs to a class of polyphenols called catechins, predominantly found in tea. The main source of EGCG is green tea, but it is also found in white, oolong, and black teas. Small amounts can also be found in berries and nuts such as pecans and hazelnuts, for example [30].
One of the interests of nutritional senolytics research is to identify other senolytics present in foods in more meaningful amounts. For example, a recent study that screened natural food compounds had the specific goal of finding new senolytics available in foods at higher amounts than fisetin [47]. The study searched for compounds that could reactivate apoptotic processes in senescent cells through a pathway fisetin also targets [inhibition of PI3K [48,49], an activator of the PI3K-AKT anti-apoptotic pathway].
The study revealed 23 natural compounds that targeted that pathway, but out of those, only five are found more abundantly in food sources: kaempferol (mentioned above), eriodictyol, cyanidin, cianidanol, ellagic acid [47]. Cianidanol is a catechin found in tea, eriodictyol can be found in citrus, and ellagic acid is found in high amounts in pomegranate, for example. Cyanidin, an anthocyanin flavonoid found in high amounts in red fruits and berries, has already shown senolytic potential by delaying the aging process in a stress-induced premature senescence cellular model [50]. But even these compounds are supplied in the diet in amounts far below what is expected to have senolytic actions in the body.
Why Are Senolytic Compounds Found in Plant Foods?
You may have noticed a pattern in the food sources of the senolytic compounds mentioned above: they are all fruits, vegetables, herbs, and spices. These are all foods that are rich in polyphenols. You may also have noticed that most of the senolytic compounds we mentioned are in fact polyphenols.
So what are polyphenols? Polyphenols are a family of natural compounds found abundantly in plants. They are divided into four main classes: phenolic acids, flavonoids, stilbenes, and lignans. Flavonoids are further divided into several subclasses including flavones, flavonols, flavanols, flavanones, isoflavones, proanthocyanidins, and anthocyanins.
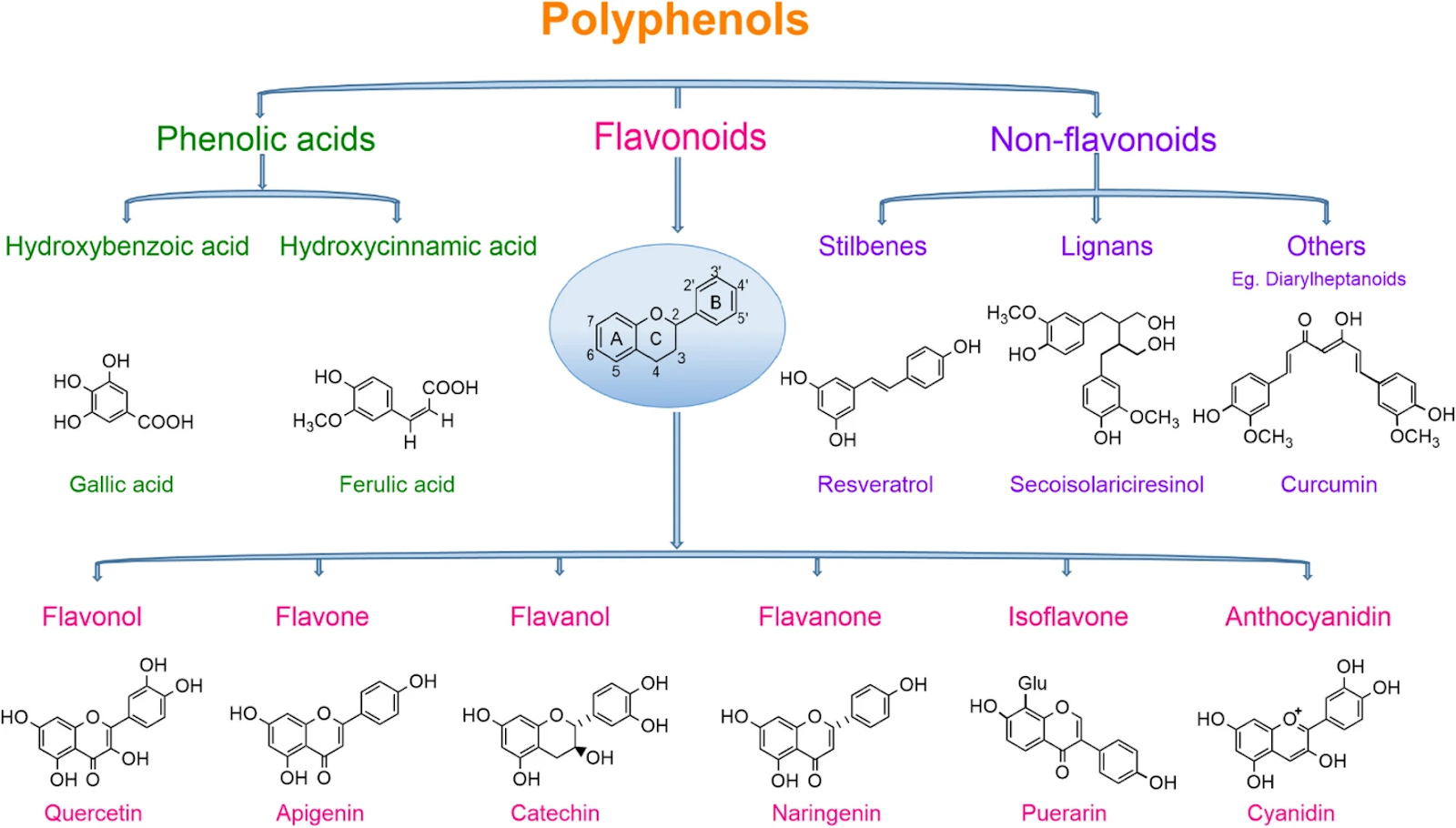
Figure 1 - Polyphenols. Source: Rambaran, TF. SN Appl. Sci. 2, 1335 (2020). License: CC BY 4.0
Polyphenols play many important roles in plant physiology. Many polyphenols are plant pigments that protect plants from environmental stressors while giving them their characteristic colors. For example, luteolin and curcumin are both yellow pigments—it’s curcumin that gives turmeric its color; anthocyanins are pigments that vary in color between red, purple, blue, and black, and which, as you may have guessed based on this color palette, are found in high quantities in berries. Because they are defense compounds, polyphenols tend to concentrate where the plant interacts with the environment. That’s why they’re often found in the highest amounts in the skins or peels of fruits and vegetables (like quercetin in onion skins).
In the human body, dietary polyphenols also support protection from stressors, both environmental and endogenous. Polyphenols support stress response and defense mechanisms such as antioxidant defenses, help to preserve healthy cell and tissue function, and support the gut microbiome, contributing to general health, including metabolic, cardiovascular, and brain health [51].
As we’ve seen, based on current knowledge from scientific studies, polyphenols are not found in foods in sufficient amounts for senolytic benefits. To get the necessary amounts from food alone would require excessive amounts of those foods. But you will still get plenty of other health benefits from eating polyphenol-rich foods.
Foods rich in polyphenols include:
- Fruits
- Vegetables
- Herbs
- Spices
- Nuts
- Algae
- Whole Cereals and Grains
- Legumes
- Cocoa
- Extra virgin olive oil
- Coffee
- Green tea and black tea
- Red wine
The way to get the studied amounts of senolytic compounds is through senolytic supplements such as Qualia Senolytic. Senolytic supplements can supply specific senolytic compounds in amounts that are unattainable through diet alone.
Qualia Senolytic
Qualia Senolytic contains a combination of nine ingredients that were carefully selected to provide a comprehensive approach to senescent cell management. All nine ingredients in Qualia Senolytic were selected because they have shown senolytic potential in preclinical research by supporting the removal of senescent cells by apoptosis or immune clearance.*
Some of the ingredients in Qualia Senolytic are being studied in ongoing clinical trials, namely fisetin and quercetin. We chose the servings of our ingredients based on those studies and on the complementarity between our different ingredients, ensuring that different mechanisms, pathways, and tissue specificities were targeted by our formulation.*
Qualia Senolytic was developed with the goal of supporting healthy aging by helping to bring the creation and clearance of senescent cells back into a healthy balance. Qualia Senolytic was also designed to support the efficient use of cellular resources, promote the growth of more youthful cells by eliminating senescent cells, support healthy tissue function, and revitalize aging tissues, promoting whole-body rejuvenation.*
Learn more about Qualia Senolytic in The Formulator's View of the Qualia Senolytic Ingredients.

References
[1]N. Herranz, J. Gil, J. Clin. Invest. 128 (2018) 1238–1246.
[2]R. Kumari, P. Jat, Front Cell Dev Biol 9 (2021) 645593.
[3]J.M. van Deursen, Nature 509 (2014) 439–446.
[4]S. Chaib, T. Tchkonia, J.L. Kirkland, Nat. Med. 28 (2022) 1556–1568.
[5]D. Muñoz-Espín, M. Serrano, Nat. Rev. Mol. Cell Biol. 15 (2014) 482–496.
[6]B.G. Childs, M. Gluscevic, D.J. Baker, R.-M. Laberge, D. Marquess, J. Dananberg, J.M. van Deursen, Nat. Rev. Drug Discov. 16 (2017) 718–735.
[7]S. He, N.E. Sharpless, Cell 169 (2017) 1000–1011.
[8]C. López-Otín, M.A. Blasco, L. Partridge, M. Serrano, G. Kroemer, Cell 186 (2023) 243–278.
[9]C.S.L. Tuttle, M.E.C. Waaijer, M.S. Slee-Valentijn, T. Stijnen, R. Westendorp, A.B. Maier, Aging Cell 19 (2020) e13083.
[10]Y. Ovadya, T. Landsberger, H. Leins, E. Vadai, H. Gal, A. Biran, R. Yosef, A. Sagiv, A. Agrawal, A. Shapira, J. Windheim, M. Tsoory, R. Schirmbeck, I. Amit, H. Geiger, V. Krizhanovsky, Nat. Commun. 9 (2018) 5435.
[11]G. Nelson, J. Wordsworth, C. Wang, D. Jurk, C. Lawless, C. Martin-Ruiz, T. von Zglinicki, Aging Cell 11 (2012) 345–349.
[12]D. McHugh, J. Gil, J. Cell Biol. 217 (2018) 65–77.
[13]N. Musi, J.M. Valentine, K.R. Sickora, E. Baeuerle, C.S. Thompson, Q. Shen, M.E. Orr, Aging Cell 17 (2018) e12840.
[14]J.N. Justice, H. Gregory, T. Tchkonia, N.K. LeBrasseur, J.L. Kirkland, S.B. Kritchevsky, B.J. Nicklas, J. Gerontol. A Biol. Sci. Med. Sci. 73 (2018) 939–945.
[15]M. Xu, T. Pirtskhalava, J.N. Farr, B.M. Weigand, A.K. Palmer, M.M. Weivoda, C.L. Inman, M.B. Ogrodnik, C.M. Hachfeld, D.G. Fraser, J.L. Onken, K.O. Johnson, G.C. Verzosa, L.G.P. Langhi, M. Weigl, N. Giorgadze, N.K. LeBrasseur, J.D. Miller, D. Jurk, R.J. Singh, D.B. Allison, K. Ejima, G.B. Hubbard, Y. Ikeno, H. Cubro, V.D. Garovic, X. Hou, S.J. Weroha, P.D. Robbins, L.J. Niedernhofer, S. Khosla, T. Tchkonia, J.L. Kirkland, Nat. Med. 24 (2018) 1246–1256.
[16]O.H. Jeon, C. Kim, R.-M. Laberge, M. Demaria, S. Rathod, A.P. Vasserot, J.W. Chung, D.H. Kim, Y. Poon, N. David, D.J. Baker, J.M. van Deursen, J. Campisi, J.H. Elisseeff, Nat. Med. 23 (2017) 775–781.
[17]J.N. Farr, M. Xu, M.M. Weivoda, D.G. Monroe, D.G. Fraser, J.L. Onken, B.A. Negley, J.G. Sfeir, M.B. Ogrodnik, C.M. Hachfeld, N.K. LeBrasseur, M.T. Drake, R.J. Pignolo, T. Pirtskhalava, T. Tchkonia, M.J. Oursler, J.L. Kirkland, S. Khosla, Nat. Med. 23 (2017) 1072–1079.
[18]E. Dookun, A. Walaszczyk, R. Redgrave, P. Palmowski, S. Tual-Chalot, A. Suwana, J. Chapman, E. Jirkovsky, L. Donastorg Sosa, E. Gill, O.E. Yausep, Y. Santin, J. Mialet-Perez, W. Andrew Owens, D. Grieve, I. Spyridopoulos, M. Taggart, H.M. Arthur, J.F. Passos, G.D. Richardson, Aging Cell 19 (2020) e13249.
[19]K.J. Mylonas, E.D. O’Sullivan, D. Humphries, D.P. Baird, M.-H. Docherty, S.A. Neely, P.J. Krimpenfort, A. Melk, R. Schmitt, S. Ferreira-Gonzalez, S.J. Forbes, J. Hughes, D.A. Ferenbach, Sci. Transl. Med. 13 (2021).
[20]T.J. Bussian, A. Aziz, C.F. Meyer, B.L. Swenson, J.M. van Deursen, D.J. Baker, Nature 562 (2018) 578–582.
[21]P. Zhang, Y. Kishimoto, I. Grammatikakis, K. Gottimukkala, R.G. Cutler, S. Zhang, K. Abdelmohsen, V.A. Bohr, J. Misra Sen, M. Gorospe, M.P. Mattson, Nat. Neurosci. 22 (2019) 719–728.
[22]S. Pathak, S. Regmi, T.T. Nguyen, B. Gupta, M. Gautam, C.S. Yong, J.O. Kim, Y. Son, J.-R. Kim, M.H. Park, Y.K. Bae, S.Y. Park, D. Jeong, S. Yook, J.-H. Jeong, Acta Biomater. 75 (2018) 287–299.
[23]M. Ogrodnik, S. Miwa, T. Tchkonia, D. Tiniakos, C.L. Wilson, A. Lahat, C.P. Day, A. Burt, A. Palmer, Q.M. Anstee, S.N. Grellscheid, J.H.J. Hoeijmakers, S. Barnhoorn, D.A. Mann, T.G. Bird, W.P. Vermeij, J.L. Kirkland, J.F. Passos, T. von Zglinicki, D. Jurk, Nat. Commun. 8 (2017) 15691.
[24]L.J. Hickson, L.G.P. Langhi Prata, S.A. Bobart, T.K. Evans, N. Giorgadze, S.K. Hashmi, S.M. Herrmann, M.D. Jensen, Q. Jia, K.L. Jordan, T.A. Kellogg, S. Khosla, D.M. Koerber, A.B. Lagnado, D.K. Lawson, N.K. LeBrasseur, L.O. Lerman, K.M. McDonald, T.J. McKenzie, J.F. Passos, R.J. Pignolo, T. Pirtskhalava, I.M. Saadiq, K.K. Schaefer, S.C. Textor, S.G. Victorelli, T.L. Volkman, A. Xue, M.A. Wentworth, E.O. Wissler Gerdes, Y. Zhu, T. Tchkonia, J.L. Kirkland, EBioMedicine 47 (2019) 446–456.
[25]J.N. Justice, A.M. Nambiar, T. Tchkonia, N.K. LeBrasseur, R. Pascual, S.K. Hashmi, L. Prata, M.M. Masternak, S.B. Kritchevsky, N. Musi, J.L. Kirkland, EBioMedicine 40 (2019) 554–563.
[26]N.S. Gasek, G.A. Kuchel, J.L. Kirkland, M. Xu, Nat. Aging 1 (2021) 870–879.
[27]Y. Zhu, T. Tchkonia, T. Pirtskhalava, A.C. Gower, H. Ding, N. Giorgadze, A.K. Palmer, Y. Ikeno, G.B. Hubbard, M. Lenburg, S.P. O’Hara, N.F. LaRusso, J.D. Miller, C.M. Roos, G.C. Verzosa, N.K. LeBrasseur, J.D. Wren, J.N. Farr, S. Khosla, M.B. Stout, S.J. McGowan, H. Fuhrmann-Stroissnigg, A.U. Gurkar, J. Zhao, D. Colangelo, A. Dorronsoro, Y.Y. Ling, A.S. Barghouthy, D.C. Navarro, T. Sano, P.D. Robbins, L.J. Niedernhofer, J.L. Kirkland, Aging Cell 14 (2015) 644–658.
[28]C. Luís, A.T. Maduro, P. Pereira, J.J. Mendes, R. Soares, R. Ramalho, Front Nutr 9 (2022) 958563.
[29]W.M. Dabeek, M.V. Marra, Nutrients 11 (2019).
[30]D.B. Haytowitz, X. Wu, S. Bhagwat, US Department of Agriculture 173 (2018).
[31]J.-H. Kwak, J.M. Seo, N.-H. Kim, M.V. Arasu, S. Kim, M.K. Yoon, S.-J. Kim, Saudi J. Biol. Sci. 24 (2017) 1387–1391.
[32]B. Santiago, A. Arias Calvo, B. Gullón, G. Feijoo, M.T. Moreira, S. González-García, Chem. Eng. J. 392 (2020) 123772.
[33]M.J. Yousefzadeh, Y. Zhu, S.J. McGowan, L. Angelini, H. Fuhrmann-Stroissnigg, M. Xu, Y.Y. Ling, K.I. Melos, T. Pirtskhalava, C.L. Inman, C. McGuckian, E.A. Wade, J.I. Kato, D. Grassi, M. Wentworth, C.E. Burd, E.A. Arriaga, W.L. Ladiges, T. Tchkonia, J.L. Kirkland, P.D. Robbins, L.J. Niedernhofer, EBioMedicine 36 (2018) 18–28.
[34]A.H. Rahmani, A. Almatroudi, K.S. Allemailem, A.A. Khan, S.A. Almatroodi, Molecules 27 (2022).
[35]M. Kimira, Y. Arai, K. Shimoi, S. Watanabe, J. Epidemiol. 8 (1998) 168–175.
[36]Y. Wang, J. Chang, X. Liu, X. Zhang, S. Zhang, X. Zhang, D. Zhou, G. Zheng, Aging 8 (2016) 2915–2926.
[37]M. Varela-Eirín, P. Carpintero-Fernández, A. Sánchez-Temprano, A. Varela-Vázquez, C.L. Paíno, A. Casado-Díaz, A.C. Continente, V. Mato, E. Fonseca, M. Kandouz, A. Blanco, J.R. Caeiro, M.D. Mayán, Aging 12 (2020) 15882–15905.
[38]G.L. Hostetler, R.A. Ralston, S.J. Schwartz, Adv. Nutr. 8 (2017) 423–435.
[39]S.J. Hewlings, D.S. Kalman, Foods 6 (2017).
[40]V. Yadav, A. Krishnan, D. Vohora, J. Ethnopharmacol. 247 (2020) 112255.
[41]S.H. Omar, Sci. Pharm. 78 (2010) 133–154.
[42]J. Burns, T. Yokota, H. Ashihara, M.E.J. Lean, A. Crozier, J. Agric. Food Chem. 50 (2002) 3337–3340.
[43]A.M. Rimando, W. Kalt, J.B. Magee, J. Dewey, J.R. Ballington, J. Agric. Food Chem. 52 (2004) 4713–4719.
[44]W.J. Hurst, J.A. Glinski, K.B. Miller, J. Apgar, M.H. Davey, D.A. Stuart, J. Agric. Food Chem. 56 (2008) 8374–8378.
[45]D.L. McKay, J.B. Blumberg, Phytother. Res. 20 (2006) 519–530.
[46]S. Shukla, S. Gupta, Pharm. Res. 27 (2010) 962–978.
[47]T. Chaganti, B. Pendyala, bioRxiv (2022) 2022.05.12.491721.
[48]C.-S. Chien, K.-H. Shen, J.-S. Huang, S.-C. Ko, Y.-W. Shih, Mol. Cell. Biochem. 333 (2010) 169–180.
[49]J.Y. Lim, J.Y. Lee, B.J. Byun, S.H. Kim, Toxicol Rep 2 (2015) 984–989.
[50]M.J. Choi, B.K. Kim, K.Y. Park, T. Yokozawa, Y.O. Song, E.J. Cho, Biol. Pharm. Bull. 33 (2010) 421–426.
[51]A. Rana, M. Samtiya, T. Dhewa, V. Mishra, R.E. Aluko, J. Food Biochem. 46 (2022) e14264.






No Comments Yet
Sign in or Register to Comment